
Bruce Edmonds (2006)
The Emergence of Symbiotic Groups Resulting from Skill-Differentiation and Tags
Journal of Artificial Societies and Social Simulation
vol. 9, no. 1
<https://www.jasss.org/9/1/10.html>
For information about citing this article, click here
Received: 12-Jun-2005 Accepted: 18-Aug-2005 Published: 31-Jan-2006

 Abstract
Abstract
|
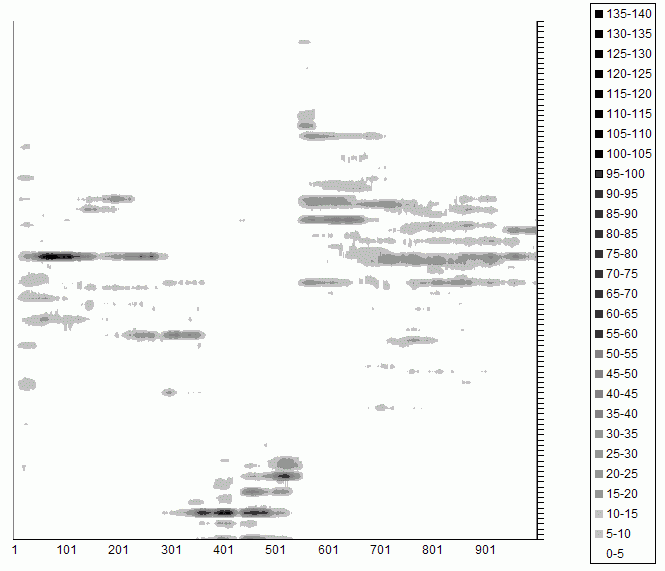
| Figure 1. Tag Groups in a Single Run of the Simulation: the vertical axis are the tag values from 0 to 1; the horizontal axis is time; the shade indicates the number of individuals with that tag at that time |
|
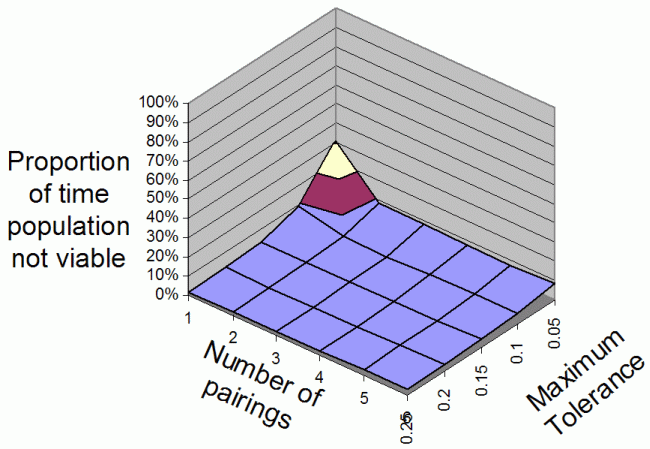
| Figure 2. The average proportion of iterations that the population is non-viable overall range of Number Of Parings and Maximum Tolerances for 2 food types |
|
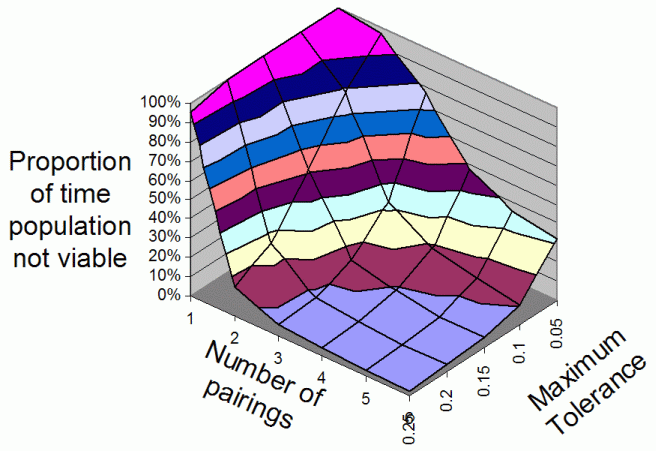
| Figure 3. The average proportion of iterations that the population is non-viable overall range of Number Of Parings and Maximum Tolerances for 3 food types |
|
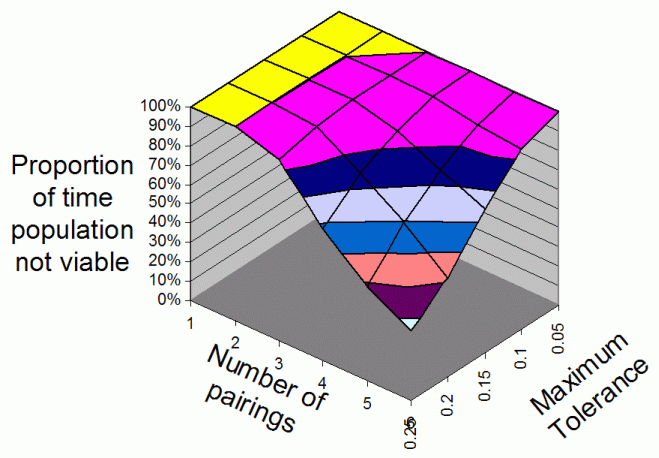
| Figure 4. The average proportion of iterations that the population is non-viable overall range of Number Of Parings and Maximum Tolerances for 4 food types |
DonationBenefit=0.95; DonationCost=1.0; MaxTime=450; FoodOfTypeAboveWhichIsExtra=5.0; FoodOfTypeBelowWhichTagDies=0.0; FoodOfTypeNecessaryForReproduction=4.0; FoodUsageRate=0.25; InitialFood=1.0; InitialPopSize=100; MaxNumNew=2; MaxReservoir=7.5; MaxStartAge=0; MaxTagAge=15; MaxTol=0.1; NumFood=200; NumFoodTypes=3; NumPairings=6; ProbMutVal=0.05; SdMut=0.05.
|
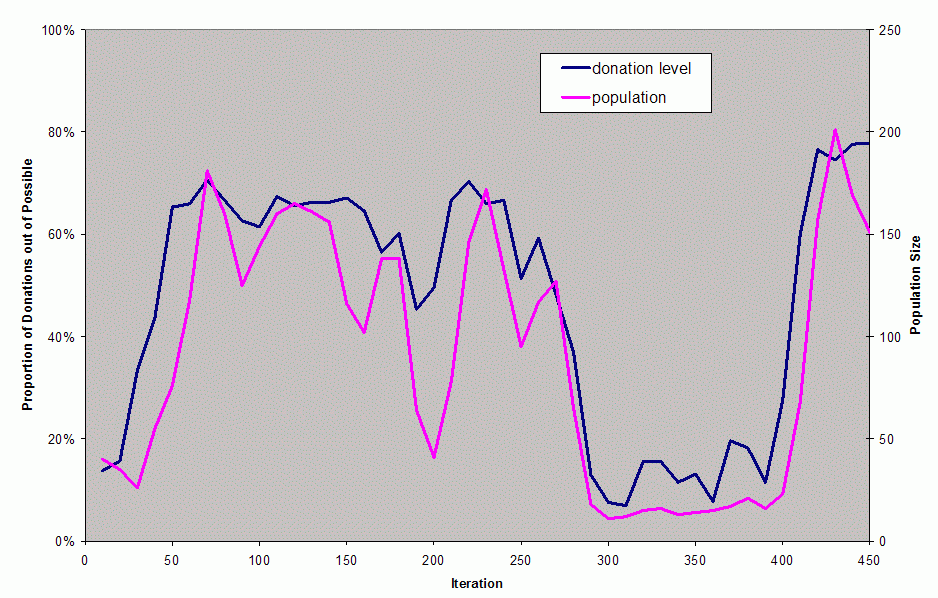
| Figure 5. The population and donation level in first 450 iteration of this simulation |
|
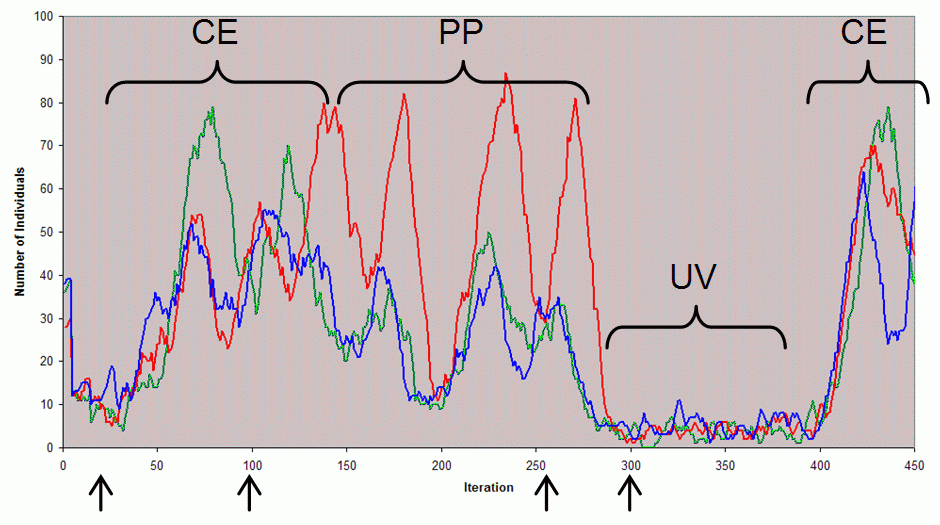
| Figure 6. The number of individuals with each of the three skills (red, green and blue) in a sample run, illustrating some of the phases that can occur. CE — coexistence: this is when there co-existing cooperative populations have been established exhibiting minor oscillations (illustrated at iteration 100 in Figure 8); PP — predator-prey: this is where parasites have arisen and thus predator-prey dynamics increasingly dominate until the population become unviable again (illustrated at iteration 260 in Figure 9); UV — unviable: this is when the population is not in a self-sustaining state (illustrated at iteration 300 in Figure 10). The arrows show the timing of snapshots of the populations that are illustrated in the following figures |
|
| Figure 7. A profile of the population at iteration 20. The population is just beginning to organise into groups with those with tag values around 0.9 and 0.1 being candidates, having all three skills and all being within the tolerance range of each other. The cluster around 0.9 will win out due to resource competition and the self-reinforcing nature of the groups in this model |
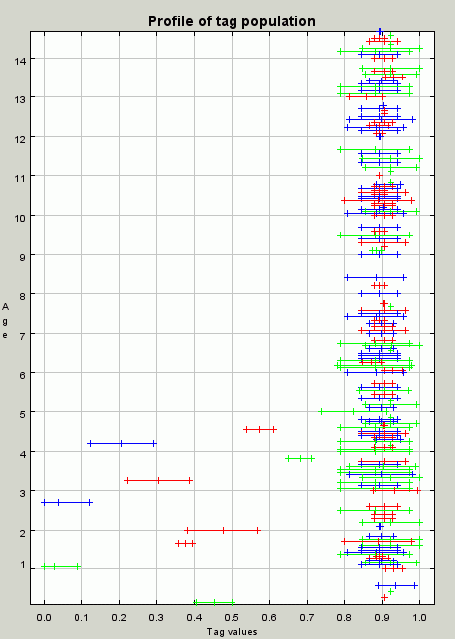
|
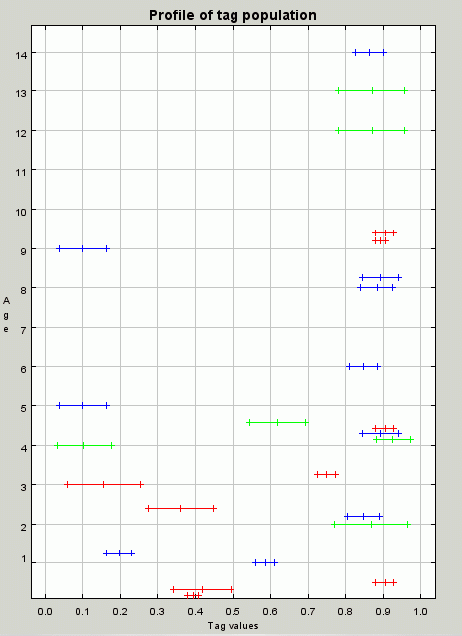
| Figure 8. A profile of the population at iteration 100. A profile of the population at iteration 100 showing the cluster of individuals with tag and tolerance values around 0.9 that allow for the effective redistribution of resources |
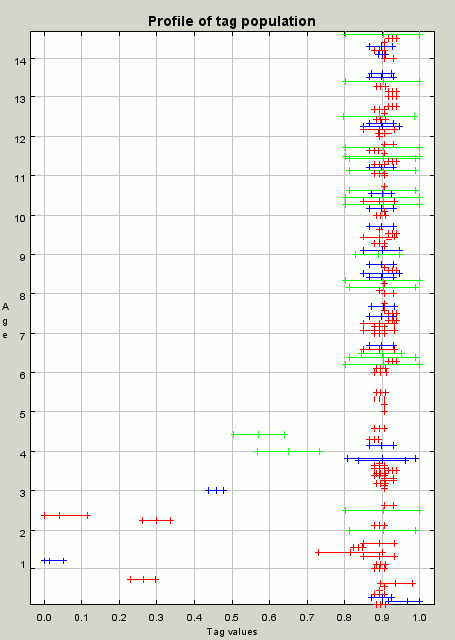
|
| Figure 9. A profile of the population at iteration 260. Although there are still cooperating individuals but there are also parasites (some of the smaller tolerance red individuals). These will soon kill of the group and themselves so that the population becomes unviable again |
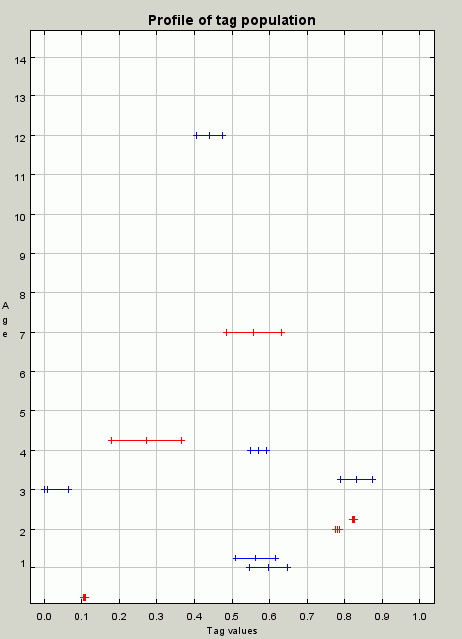
|
| Figure 10. A profile of the population at iteration 300. There are no skills left of the green type and so the population is unviable |
A QuickTime movie of the profiles in this run, every 10 iterations, is available here.
2The individuals are too simple to be called 'agents', though happen to be simulated as separate objects with rules. One can well imagine their dynamics occurring in creatures without anything we would call cognition.
3In other tag models different ways of representing tags have been used (including multi-dimensional binary and continuous vectors and integers) — in the models that have been tried this does not seem to significantly change the results.
4It is important that the relation is strict ('<') rather than not ('≤') as this allows for totally selfish individuals to exist — otherwise (as documented in Edmonds and Hales 2003) individuals have no option but to donate to clones of themselves.
5Indeed, the self-creation and regulation of "group" identity here might allow one to claim that the groups are autopoetic (Maturana and Valera 1980).
6The existence of emergence, such as observed in these cases, calls into question the assumption that a finer level of description has any sort of precedence over descriptions at higher levels. What is of more importance are the properties of persistence, continuity, usefulness, identifiably, predictability etc. As Anderson (1972) put it: "more is different".
7A review of the literature on group selection is Wilson and Sober (1994)
8For background on these ideas see Barkow et al. (1992).
Generate individuals, giving them all initialFood in each resource
and each an independent randomly chosen skill, tag and tolerance
For each generation
Add maxNumNew new individuals (with random tags)
Units of resource are randomly distributed among nutrition types -
individuals with a skill equally share in that type
For each individual, D
For each pairing from 1 to numPairings
Randomly choose another individual without replacement, O
For each resource type, R
If
D has more of R than foodOfTypeAboveWhichIsExtra
and the absolute difference between D's and O's tag
is strictly less than D's tolerance
Then
Subtract donationCost in R from D
Add donationBenefit in R to O
Next resource type
Next pairing
Next individual
For each individual
subtract foodUsageRate from each resource
If any resource < foodOfTypeBelowWhichTagDies then it dies
If
all resources > foodOfTypeNecessaryForReproduction
Then
replicate individual (with possible mutation),
subtracting new progeny's resources from parent
Next individual
Next generation
|
| Figure A1. Algorithm |
AXELROD, R. (1984) The Evolution of Cooperation, Basic Books, New York.
BARKOW, J. H., Cosmides, L. & Tooby, J. (Eds.), (1992) The adapted mind: evolutionary psychology and the generation of culture. Oxford University Press.
EDMONDS, B. and Hales, D. (2003) Replication, Replication and Replication - Some Hard Lessons from Model Alignment. Journal of Artificial Societies and Social Simulation 6(4). https://www.jasss.org/6/4/11.html
EDMONDS, B. and Hales, D. (2005) Computational Simulation as Theoretical Experiment, Journal of Mathematical Sociology, 29:1-24.
HALES, D. (2000) Cooperation without Space or Memory: Tags, Groups and the Prisoner's Dilemma. In Moss & Davidsson, (eds.) Multi-Agent-Based Simulation. LNAI 1979:157-166. Springer. Berlin, 2000.
HALES, D. (2001) Tag Based Co-operation in Artificial Societies. PhD Thesis, Department of Computer Science, University of Essex, UK, 2001.
HALES, D. (2004) Understanding Tag Systems by Comparing Tag Models. Second Model-to-Model Workshop (M2M2) at the Second European Social Simulation Association Conference (ESSA'04) Valladolid, Spain 16-19th Sept 2004.
HALES, D. (2005) Choose Your Tribe! - Evolution at the Next Level in a Peer-to-Peer Network. University of Bologna, Dept. of Computer Science, May 2005, Technical Report UBLCS-2005-13. http://www.cs.unibo.it/pub/TR/UBLCS/ABSTRACTS/2005.bib
HALES, D. and Edmonds, B. (2005) Applying a socially-inspired technique (tags) to improve cooperation in P2P Networks, IEEE Transactions in Systems, Man and Cybernetics, 35:385-395.
HARDIN, G. (1968) The Tragedy of the Commons, Science, 162:1243-1248.
HOLLAND, J. (1993) The Effect of Labels (Tags) on Social Interactions. SFI Working Paper 93 10 064. Santa Fe Institute, Santa Fe, New Mexico, USA.
HOLLAND, J. (1995) Hidden order : how adaptation builds complexity Reading, MA: Addison-Wesley.
KUMMER, H., Daston, L., Gigerenzer, G. & Silk, J. (1997) The social intelligence hypothesis. In Weingart et. al (eds.), Human by Nature: between biology and the social sciences. Hillsdale, NJ: Lawrence Erlbaum Associates, 157-179.
MARGULIS, L. and Sagan, D. Microcosmos: Four Billion Years of Evolution from Our Microbial Ancestors. Summit Books, New York, 1986.
MATURANA, H. R. and Varela, F. J. (1980) Autopoiesis and cognition : the realization of the living. Boston studies in the philosophy of science; v.42, Kluwer Academic.
READER, J. Man on Earth. London: Collins, 1988.
RIOLO, R. (1997). The effects of tag-mediated selection of partners in evolving populations playing the iterated Prisoner's Dilemma. Santa Fe Institute Working Paper, 97-02-016. http://www.cscs.umich.edu/pub/papers/icga97-ipd.ps
RIOLO, R. L., Cohen, M. D. and Axelrod, R. (2001) Evolution of cooperation without reciprocity. Nature, 411:441-443.
ROBERTS, G. and Sherratt, T. (2002) Does similarity breed cooperation? Nature 418:499-500.
SIGMUND, K. and Nowak, M. A. (2001) Evolution - Tides of tolerance. Nature 414:403.
TAKAHASHI, N. (2000) The Emergence of Generalized Exchange. American Journal of Sociology 105:1105-1134,.
WILSON, D.S. & Sober, E. (1994) Reintroducing group selection to the human behavioural sciences. Behavioral and Brain Sciences 17(4): 585-654.
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, [2006]