
Stephen Younger (2005)
Violence and Revenge in Egalitarian Societies
Journal of Artificial Societies and Social Simulation
vol. 8, no. 4
<https://www.jasss.org/8/4/11.html>
For information about citing this article, click here
Received: 04-May-2005 Accepted: 08-Aug-2005 Published: 31-Oct-2005

 Abstract
Abstract
| Altruism parameter, Ai: | 1/3 chance of inheriting Amother - 0.1 |
| 1/3 chance of inheriting Amother | |
| 1/3 chance of inheriting Amother + 0.1 | |
| Aggression parameter, Gi: | Random assignment |
| Fighting ability, Fi: | Random assignment |
The scheme chosen for the altruism parameter, i.e. modified matrilineal inheritance of the normative character of the mother, represented both the effect of genetic inheritance and the nurturing of the young by the mother. Some variation was introduced into this inheritance, a form of genetic noise. A discussion of the effect of the inheritance of normative characteristics using a simulation method similar to the one used here can be found in Younger (2005a).
| Table 1: Results comparing the cases of no violence, violence only, and violence plus revenge. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | |||
| No Violence Or Revenge | Violence, but No Revenge | Both Violence and Revenge | |
| Survival rate (%) | 100 | 45 | 35 |
| Deaths due to old age (%) | 72 | 72 | 70 |
| Deaths due to hunger (%) | 28 | 6 | 4 |
| Deaths due to violence (%) | 0 | 22 | 12 |
| Deaths due to revenge (%) | 0 | 0 | 14 |
| Total Mutual Obligation | 360 | 410 | 380 |
| Table 2: Results of excluding different segments of the population from violence. "Family" means that violence was forbidden within the family, "Group" within the group, etc. The last column represents a situation where violence was permitted only against males of the other group. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | ||||||||
| Subpopulation excluded from violence | None | Females | Group | Family | Group Females | Family Females | Family Group | Family Group Females |
| Survival rate (%) | 45 | 85 | 65 | 30 | 100 | 80 | 55 | 90 |
| Deaths due to old age (%) | 72 | 70 | 67 | 74 | 70 | 69 | 72 | 68 |
| Deaths due to hunger (%) | 6 | 21 | 13 | 9 | 23 | 24 | 18 | 27 |
| Deaths due to violence (%) | 22 | 9 | 19 | 17 | 7 | 7 | 10 | 5 |
| Total Mutual Obligation | 410 | 340 | 410 | 410 | 350 | 340 | 420 | 360 |
| Table 3: Results of excluding different segments of the population from violence and revenge. "Family" means that violence and revenge were forbidden within the family, "Group" within the group, etc. The last column represents a situation where violence and revenge were permitted only against males of the other group. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | ||||||||
Subpopulation excluded from violence and revenge | None | Females | Group | Family | Group Females | Family Females | Family Group | Family Group Females |
| Survival rate (%) | 35 | 40 | 30 | 10 | 60 | 35 | 55 | 90 |
| Deaths due to old age (%) | 70 | 75 | 68 | 71 | 71 | 71 | 71 | 74 |
| Deaths due to hunger (%) | 4 | 13 | 4 | 3 | 16 | 17 | 18 | 19 |
| Deaths due to violence (%) | 12 | 3 | 11 | 8 | 3 | 3 | 4 | 2 |
| Deaths due to revenge (%) | 14 | 9 | 17 | 19 | 10 | 9 | 7 | 5 |
| Total Mutual Obligation | 380 | 330 | 400 | 420 | 340 | 310 | 430 | 370 |
|
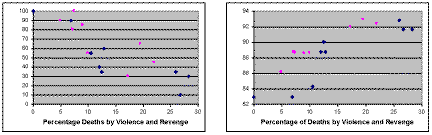
| Figure 1. Left: Survival rate of total population vs. percentage of deaths due to violence and revenge. Right: Percentage of agents with Ai > 0.5, i.e. those more likely to share, vs. percentage of deaths due to violence and revenge. Blue diamonds: Violence and revenge. Purple squares: Violence but no revenge |
|
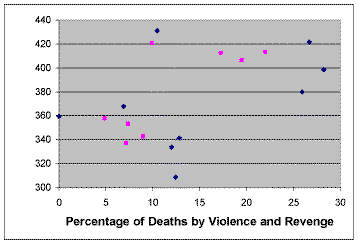
| Figure 2. Total mutual obligation vs. percentage of deaths due to violence and revenge. Blue diamonds: Violence and revenge. Purple squares: Violence but no revenge |
| Table 4: Effect of permitting varying degrees of tolerance before revenge was invoked. In this case violence and revenge were permitted between members of a group and between members of different groups, but not among family members or on females. The last row refers to the case where the revenge was not permitted. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | ||||||
| Tolerance | Survival Rate (%) | Deaths Age (%) | Deaths Hunger (%) | Deaths Violence (%) | Deaths Revenge (%) | Mutual Obligation |
| 0 | 35 | 71 | 17 | 3 | 9 | 310 |
| -25 | 70 | 73 | 16 | 6 | 5 | 350 |
| -50 | 60 | 69 | 22 | 6 | 3 | 360 |
| -101 | 65 | 74 | 16 | 7 | 3 | 360 |
| -200 | 75 | 75 | 16 | 6 | 3 | 360 |
| -400 | 85 | 71 | 22 | 7 | 0.8 | 370 |
| -800 | 55 | 69 | 22 | 8 | 0.7 | 320 |
| -1600 | 80 | 69 | 23 | 7 | 1 | 340 |
| -3200 | 70 | 69 | 23 | 7 | 0.6 | 330 |
| -6400 | 80 | 68 | 23 | 8 | 0 | 330 |
| -12800 | 70 | 69 | 24 | 7 | 0 | 330 |
| No Rev | 80 | 69 | 24 | 7 | 0 | 340 |
|
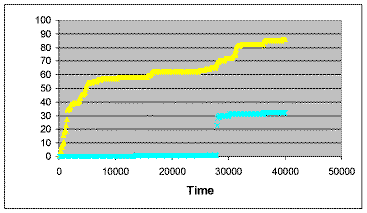
| Figure 3. Time history of cumulative deaths due to violence and revenge for one run where the tolerance to revenge was — 3200. Yellow triangles: Violence. Blue crosses: Revenge |
| Table 5: Comparison of runs with maximum sustainable populations of 100 agents and 400 agents in the scenario where violence and revenge were permitted between members of a group and between members of different groups but not among family members or among females. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | ||
| Sustainable population | 100 | 400 |
| Survival rate (%) | 35 | 70 |
| Deaths due to old age (%) | 71 | 78 |
| Deaths due to hunger (%) | 17 | 4 |
| Deaths due to violence (%) | 3 | 3 |
| Deaths due to revenge (%) | 9 | 15 |
| Mutual Obligation | 310 | 420 |
| Table 6: Relative contributions to mortality of age, hunger, violence, and revenge for a linearly decreasing food supply. Constant Food Nonzero refers to runs that had a non-zero ending population in a constant food landscape. Constant Food Zero refers to runs that had a zero ending population in a constant food landscape, i.e. unsuccessful populations in a food-rich environment. Decreasing Food refers to runs conducted with a linearly decreasing food replenishment rate. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | |||
| Constant Food Nonzero | Constant Food Zero | Decreasing Food Zero | |
| Deaths due to old age (%) | 71 | 59 | 23 |
| Deaths due to hunger (%) | 17 | 18 | 56 |
| Deaths due to violence (%) | 3 | 6 | 5 |
| Deaths due to revenge (%) | 9 | 17 | 17 |
| Table 7: Results of simulations that allowed agents to flee from other agents with whom they had a negative interaction matrix element or an interaction matrix element below that which would result from a single directly experienced theft. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | |||
| No flight | Flight | Flight when Imx < -101 | |
| Survival rate (%) | 35 | 95 | 50 |
| Deaths due to old age (%) | 71 | 37 | 68 |
| Deaths due to hunger (%) | 17 | 45 | 17 |
| Deaths due to violence (%) | 3 | 4 | 4 |
| Deaths due to revenge (%) | 9 | 14 | 12 |
| Mutual Obligation | 310 | -267 | 112 |
| Table 8: Comparison of runs where killing reduced and where it added to the interaction matrix element connecting killer and observer in the scenario where violence and revenge were permitted between members of a group and between members of different groups but not among family members or among females. Each entry represents an average over twenty runs where only those runs that had a non-zero population at the end of the run were included in the average | ||
| Modification of interaction matrix element for killing | -4000 | +4000 |
| Survival rate (%) | 35 | 80 |
| Deaths due to old age (%) | 71 | 72 |
| Deaths due to hunger (%) | 17 | 21 |
| Deaths due to violence (%) | 3 | 4 |
| Deaths due to revenge (%) | 9 | 3 |
| Mutual Obligation | 310 | 480 |
| Table 9: Comparison of simulations with ethnographic observations | ||
| Simulation Result | Ethnographic Observations | Comment |
| Violence and revenge contributed substantially to mortality and reduced the overall survival rate of the population. | Violent deaths accounted for 10's of percent of the total recorded deaths among the Copper Eskimos, Gibusi, Waorani, and other indigenous peoples. | Violence is a population control mechanism in some egalitarian societies. |
| Excluding significant segments of the population from violence and revenge improved the survival rate of the total population. | Kapauku excluded females from violence. Little violence among females in Kumaimaipa society. Some primate and human societies proscribe violence within immediate social group. | Many societies discourage violence among significant parts of the population. |
| Tolerance before revenge increased the survival rate of the total population. | Peaceful societies (e.g. Semai) have high levels of tolerance. Violent societies (e.g. Niue) have low levels of tolerance. | Tolerance reduces the rate of revenge killing. |
| Increasing the population density increased the survival rate of the total population, even though revenge killings increased. | Keeley (1996) found that population density and the rate of violence were not correlated. | Higher survival rate in simulations a result of more mating opportunities. Simulations omit control mechanisms that limit violence in real societies. |
| The rate of violence increased when food scarcity was introduced but many more agents died of hunger than by violence and revenge. | Scarcity reduced sharing within a group and, in extreme circumstances, increased antagonism and theft within the group. (Dirks 1980). The Ik of Uganda are a particular example of theft increasing in times of scarcity. | Scarcity of food increased the rate of violence, consistent with ethnographic observations. |
| Flight from known aggressors enhanced the survival of the total population, at the expense of sharing-generated social cohesion. | Flight was common among the Buid and Semai, where violence was discouraged. It was rare among the Ilongot and Yanomomo where violence was esteemed. | Role of flight depends on the cultural values of the society. Other factors, e.g. kinship, maintain social cohesion in real societies |
| When killing had a positive social value the survival rate of the total population increased as the number of revenge killings decreased. | Yanomomo and Waorani were violent people and yet survived over many generations. | Rates of violence and revenge can be very high when they are seen as a positive social value and do not inhibit mate selection and procreation. |
Main Agent Decision Routine:
FOR each agent:
IF agent is female, has no mate, and has reached reproductive age THEN
SELECT male with whom female has highest interaction matrix element
PLACE mated female at same location as her mate
IF hunger > 0.5 maximum hunger THEN
CALL food routine
ELSE
CALL movement routine to explore landscape
END IF
CALL communications routine
END agent decision routine
Food Routine:
IF the agent is carrying food THEN eat all of that food
IF the agent is at a food source THEN
Eat food to reduce hunger to zero
IF food remains at the food source, collect up to maximum carry capacity
EXIT food routine
ELSE
SEARCH memory of agent to find nearest food source with food at time of last observation
IF agent remembers a food source with food THEN
MOVE one square in the direction of that food source
ELSE
CALL movement routine to move one square in a random direction in search of food
END IF
END IF
END food routine
Movement routine:
SENSE the environment by storing the location of all agents and food sources within sensory range
STORE the locations of all food sources, along with the amount of food and the time of observation, in the agent's memory
IF the flight option is enabled AND there is an agent within sensory range for whom the active agent has a negative interaction matrix element THEN move one square away from that agent AND EXIT the movement routine
IF the agent is searching for food AND the agent is at a food source THEN
Eat food to reduce hunger to zero
IF food remains at the food source, collect up to maximum carry capacity
EXIT the movement routine
ELSE
IF agent has sensed a food source that has food THEN
MOVE one square in the direction of that food source
ELSE
MOVE one square in a random direction in search of a food source that is in supply
END IF
END IF
Communication routine:
Sharing
IF there are other agents at the same location AND the active agent is carrying food AND the ratio (agent hunger / maximum hunger) < the agent's altruism parameter THEN
COUNT the number of agents at that location who are not carrying food
CALCULATE a share as the amount of food carried by the active agent divided by the number of counted agents
GIVE a full share to agents of the same group as the sharer and one half share to agents of a different group
ADD the amount shared to the interaction matrix element imx (receiver, sharer) of each agent
END IF
Stealing
IF there is another agent at the same location who is carrying food AND the active agent is not carrying food AND [(agent hunger / maximum hunger before death) > the agent's altruism parameter] AND [(agent hunger / maximum hunger) > ( 1 - agent's aggression parameter)] THEN
IF active agent has larger aggression parameter AND larger fighting ability parameter THEN
TAKE all of the food carried by the other agent
REDUCE the interaction matrix element imx(victim, thief) by the amount of food stolen
ELSE
IF violence is permitted between this pair of agents AND the active agent has a larger aggression parameter AND the other agent has a larger fighting ability parameter THEN
The active agent is killed
DEDUCT an amount equal to an agent lifetime from the interaction matrix elements imx(collocated, killer) for all agents who were of the same group as the agent killed and who were collocated with the killing
EXIT communication routine
ELSE
IF active agent has larger fighting ability parameter and a smaller aggression parameter than the other agent THEN
The other agent is killed
The food carried by the other agent is taken by the active agent
DEDUCT an amount equal to an agent lifetime from the interaction matrix elements imx(collocated, killer) for all agents who were of the same group as the agent killed who were collocated with the killing
END IF
END IF
END IF
Revenge
FOR all other agents at the same location as the active agent
IF the interaction matrix element imx(active agent, other agent) < 0 THEN
IF the active agent has a higher fighting ability parameter than the other agent THEN
The other agent is killed
DEDUCT an amount equal to an agent lifetime from the interaction matrix elements imx(collocated, killer) for all agents who were of the same group as the agent killed who were collocated with the killing
ELSE
The active agent is killed
DEDUCT an amount equal to an agent lifetime from the interaction matrix elements imx(collocated, killer) for all agents who were of the same group as the agent killed who were collocated with the killing
END IF
END IF
NEXT AGENT
Share normative reputation of other agents
FOR all other agents j at the same location at the active agent
FOR all agents k
IF either the active agent or agent j has a non-zero interaction matrix element imx(agent, k) THEN
imx(active, k) = 1/2 (imx(active, k) + imx(j, k))
imx(other, k) = 1/2 (imx(active, k) + imx(j, k))
NEXT AGENT k
NEXT AGENT j
Agent reproduction
IF the active agent is collocated with its mate
AND 0.25 * (agent lifetime) < male age < 0.75 * (agent lifetime)
AND 0.25 * (agent lifetime) < female age < 0.75 * (agent lifetime)
AND (RANDOM NUMBER) < 0.0045
THEN
SET parameters of a new agent
Location is same as parents
Group is same as parents
Equal probability of male or female
Random aggression parameter
Random fighting ability parameter
Altruism parameter:
1/3 chance of mother's altruism parameter
1/3 chance of mother's altruism parameter + 0.1
1/3 chance of mother's altruism parameter — 0.1
END IF
BOEHM C (1999) Hierarchy in the Forest: The Evolution of Egalitarian Behavior. Cambridge, MA: Harvard University Press.
CAMPBELL I C (1989) A History of the Pacific Islands. Berkeley, CA: University of California Press.
CAMPBELL J (1975) On the Conflicts between Biological and Social Evolution and between Psychology and Moral Tradition. American Psychologist, 30. pp. 1103-1126.
CHAGNON, N A (1983) Yanomomo: The Fierce People. New York: Holt Reinhart and Winston.
CHAGNON, N A (1990) "Reproductive and Somatic Conflicts of Interest in the Genesis of Violence and Warfare Among Tribesmen". In Hass J (Ed.), The Anthropology of War, Cambridge: Cambridge University Press.
DIRKS R (1980) Social Responses During Severe Food Shortages and Famine. Current Anthropology, 21. pp. 21-44.
FERGUSON R B (2005) "Archaeology, Cultural Anthropology, and the Origins and Intensifications of War". In Arkush E and Allen M (Eds.), Violent Transformations: The Archaeology of Warfare and Long-Term Social Change, Salt Lake City, UT: University of Utah Press.
GOLDMAN I (1970) Ancient Polynesian Society. Chicago, IL: University of Chicago Press.
GIBSON T (1990) "Raiding, Trading, and Tribal Autonomy in Insular Southeast Asia". In Hass J (Ed.), The Anthropology of War, Cambridge: Cambridge University Press.
GREGOR T (1990) "Uneasy Peace: Intertribal Relations in Brazil's Upper Xingu". In Hass J (Ed.), The Anthropology of War, Cambridge: Cambridge University Press.
HAYDEN B (1972) Population Control Among Hunter Gatherers. World Archeology, 4. pp. 205-221.
JAGER W, Popping R, and van de Sande H (2001) Clustering and Fighting in Two-party Crowds: Simulating the Approach-avoidance Conflict. Journal of Artificial Societies and Social Simulation 4(3) https://www.jasss.org/4/3/7.html.
JAFFE K (2002) An Economic Analysis of Altruism: Who Benefits from Altruistic Acts? Journal of Artificial Societies and Social Simulation 5(3) https://www.jasss.org/5/3/3.html.
KEELEY L H (1996) War Before Civilization: The Myth of the Peaceful Savage. Oxford: Oxford University Press.
KELLY R L (1995) The Foraging Spectrum: Diversity in Hunter-Gatherer Lifeways. Washington DC: Smithsonian Institution Press.
KNAUFT B M (1987) Reconsidering Violence in Simple Human Societies: Homicide Among the Gebusi of New Guinea. Current Anthropology, 28. pp. 457-500.
KNAUFT BM (1991) Violence and Sociality in Human Evolution. Current Anthropology, 32. pp. 391-428.
LEE R B (1979) The !Kung San: Men, Women, and Work in a Foraging Society. New York: Cambridge University Press.
MALTHUS T R (1798) Essay on Population
MCARTHUR A M (2000) The Curbing of Anarchy in Kunimaipa Society. Sydney: University of Sydney.
MCCAULEY C (1990) "Conference Overview". In Hass J (Ed.), The Anthropology of War, Cambridge: Cambridge University Press.
OTTERBEIN K F and Otterbein C S (1965) A Eye for an Eye, A Tooth for a Tooth: A Cross-Cultural Study of Feuding. American Anthropologist, New Series, 67. pp. 1470-1482.
OTTERBEIN K F (2000) Killing of Captured Enemies: A Cross Cultural Study. American Anthropologist, 41. pp. 439-443.
PETERSON N (1993) Demand Sharing: Reciprocity and the Pressure for Generosity Among Foragers. American Anthropologist, New Series, 95. pp. 860-874.
ROBARCHEK C and Denton R K (1987) Blood Drunkenness and the Bloodthirsty Semai: Unmaking Another Anthropological Myth. American Anthropologist, 89. pp. 356-365.
ROBARCHEK C (1990) Motivations and Material Cause: On the Explanation of Conflict and War. In Hass J (Ed.), The Anthropology of War, Cambridge: Cambridge University Press.
ROBARCHEK C A and Robarchek C J (1992) "Cultures of War and Peace: A Comparitive Study of Waorani and Semai". In Silverberg J and Gray J P (Eds.), Aggression and Peacefulness in Humans and Other Primates. New York: Oxford University Press.
ROSENFELD R and Messner S F (1991) The Social Sources of Homicide in Different Types of Society. Sociological Forum, 6. pp. 51-70.
TURNBULL C (1987) The Mountain People. New York: Touchstone.
WARNER W L (1937) A Black Civilization. New York: Harper.
WOODBURN J (1982) Egalitarian Societies. Man, 17. pp. 431-451.
YOST JA and (1981) "Twenty Years of Contact: The Mechanisms of Change in Huao (Auca) Culture". In Whitten N A (Ed.), Cultural Transformations and Ethnicity in Modern Ecuador. Urbana, IL: University of Illinois Press.
YOUNGER S M (2005) Reciprocity, Sanctions, and the Development of Mutual Obligation in Egalitarian Societies. Journal of Artificial Societies and Social Simulation 8(2) https://www.jasss.org/8/2/9.html.
YOUNGER S M (2005a) Behavioral Norms and the Evolution of Societies, Proceedings of the AISB Conference 2005 (In press).
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, [2005]