
Douglas R. White (1999)
Journal of Artificial Societies and Social Simulation vol. 2, no. 3, <https://www.jasss.org/2/3/5.html>
To cite articles published in the Journal of Artificial Societies and Social Simulation, please reference the above information and include paragraph numbers if necessary
Received: 7-Mar-99 Accepted: 5-Oct-99 Published: 31-Oct-99
![]()
 Abstract
Abstract
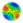
|
| Figure 1: Illustration of a "Simple" Partially ordered uniform marriage structure |
| Table 1: Normalized Endogamy (diagonal) a = ad/bc, endogamy ratio | ||
Marry: | women of A | women of B |
| Men of A | a | b |
| Men of B | c | d |
| Table 2: Normalized Alliance (off-diagonal), a = ad/bc, endogamy ratio, 1/a is the exogamy ratio | ||
Marry: | women of A | women of B |
| men of X | a | b |
| men of Y | c | d |
| Table 3: Simulated Normalization, a* = adfg / bceh, endogamy ratio | ||||||||
| Actual data | Simulated data | |||||||
| a | b | e | f | |||||
| c | d | g | h | |||||
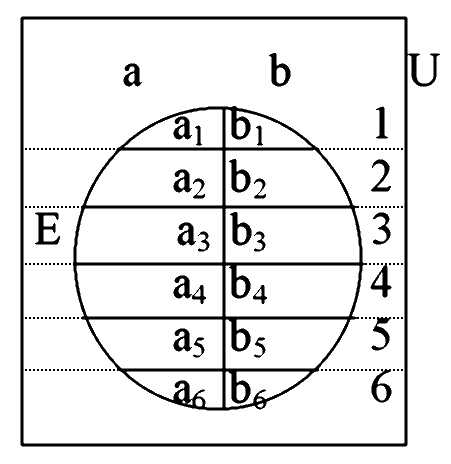
|
| Figure 2: Components, bicomponents and tricomponents of graphs, illustrating structural endogamy in graphs where marriages are the nodes and persons connecting marriages via parent-child links are the lines (direction of lines ignored) |
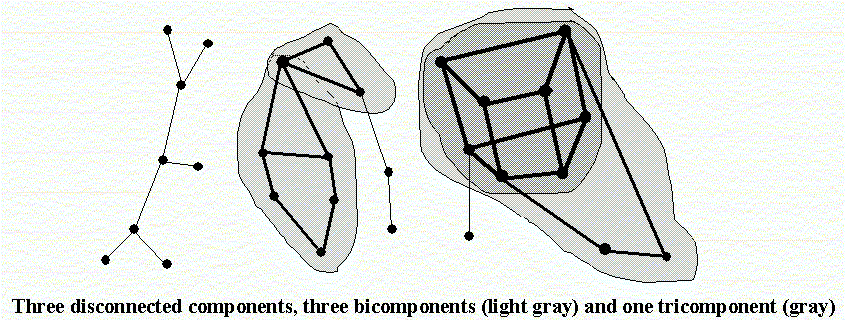
|
| Figure 3: Illustration of permuting male links in a p-graph with 3-generations: left is the origianl graph, and right is the permuted graph. Top row is the first generation of marriages, solid arrows represent sons descending from parental marriages to form a marriage with a wife, the latter shown as a broken arrow descending from her parents. |
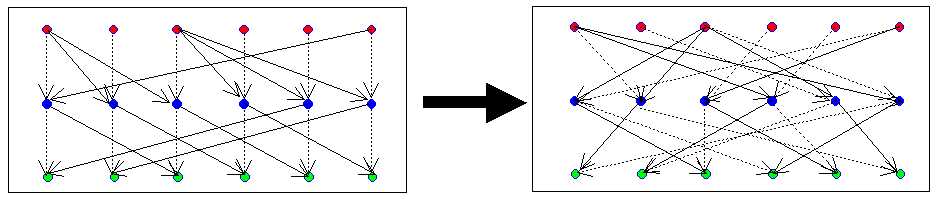
|
| Figure 4: Illustration of permuting male and female links in a p-graph with 3-generations: left is the original graph, and right is the permuted graph |
| Table 4: Case Studies and their Characteristics | ||||
| Case Studies: | 1A | 1B | 2 | 3 |
| Village | Dukuh hamlet | Dukuh Elites | Pul Eliya | Feistritz |
| Country | Indonesia | Indonesia | Sri Lanka | Austria |
| Religion | Muslim/Hindu | Muslim | Hindu-Dravidian | Catholic |
| Descent | Bilateral | Bilateral | Bilateral | Bilateral |
| Residence | Dispersed | Clustered | Diga (patrilocal v. Binna (uxorilocal) | Stem |
| Inheritance | Equal Division, 2:1 male/female | Equal Division, 2:1 male/female | Agnatic | Impartible farmsteads |
| Class/Caste | Elites/commoners | Elites | Varna subcaste with | Heir/nonheir social class |
| Marriage Structure | Status endogamy | Status endogamy | Cognatic dual (sided) organization | Class position defined by matrimonial relinking |
| Known Incest Prohibitions | Brother-sister | Brother-sister | Brother-sister | Brother-sister and 1st, 2nd cousins |
| Table 5: Comparison of Relinking Frequencies for Actual and Simulated Data (*=greater than chance), for (A) Dukuh hamlet, Indonesia (Schweizer 1989), and (B) Muslim elite of the Village containing Dukuh Hamlet (White and Schweizer 1998) | ||||||
| Magnitude of Structural Endogamy with ancestors back 1, 2, ..., g generations | ||||||
| Back: | 1 | 2 | 3 | 4 | 5 | 6 |
| (A) Dukuh | ||||||
| Starting from: | ||||||
| Present generation | ||||||
| Actual | 0 | 27 | 43 | 45 | 45 | |
| Simulated | 0 | 32 | 42 | 42 | 42 | |
| Back one generation | ||||||
| Actual | 6* | 6 | 17 | 17 | ||
| Simulated | 0 | 14 | 14 | 14 | ||
| (B) Elites | ||||||
| Starting from: | ||||||
| Present generation | ||||||
| Actual | 0 | 4 | 9 | 10 | 10 | |
| Simulated | 0 | 4 | 9 | 10 | 10 | |
| Back one generation | ||||||
| Actual | 0 | 7 | 8 | 8 | ||
| Simulated | 0 | 7 | 8 | 8 | ||
| Table 6: Comparison of Relinking Frequencies for Actual and Simulated Data (*=greater than chance) for Pul Eliya (Houseman and White 1998) | ||||||||
| Magnitude of Structural Endogamy with ancestors back 1, 2, ..., g generations | ||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Pul Eliya | ||||||||
| Starting from: | ||||||||
| Present generation | ||||||||
| Actual | 0 | 34* | 55* | 62 | 73 | 81 | 94 | 94 |
| Simulated | 0 | 26 | 48 | 63 | 71 | 84 | 93 | 93 |
| Back one generation | ||||||||
| Actual | 8* | 30* | 33 | 45 | 54 | 67 | 67 | |
| Simulated | 0 | 15 | 40 | 46 | 60 | 69 | 69 | |
| Back two generations | ||||||||
| Actual | 4* | 7* | 7* | 38 | 50 | 50 | ||
| Simulated | 0 | 0 | 0 | 37 | 46 | 46 | ||
| Back three generations | ||||||||
| Actual | 0 | 0 | 4* | 25 | 25 | |||
| Simulated | 0 | 0 | 0 | 29 | 29 | |||
| Back four generations | ||||||||
| Actual | 0 | 4* | 15 | 15 | ||||
| Simulated | 0 | 0 | 17 | 17 | ||||
| Back five generations | ||||||||
| Actual | 0 | 5 | 5 | |||||
| Simulated | 0 | 10 | 10 | |||||
| Table 7: Test of Actual versus Simulated Marriage among Consanguineal Kin, Dukuh Hamlet and Village Elites (conclusion: no preferred marriages, only status endogamy) | ||||||||||||||
| Javanese elites | Dukuh Hamlet | 3-way | ||||||||||||
| A | S | TA | TS | p= | type | A | S | TA | TS | p= | type | Test | ||
| 1: | 1 | 0 | 4 | 3 | .625 | FBD | 0 | 1 | 9 | 12 | .591 | FBD | 1.00 | |
| 2: | 1 | 2 | 2 | 3 | .714 | MBD | 1 | 0 | 11 | 16 | .429 | MBD | 1.00 | |
| 3: | 2 | 1 | 3 | 2 | .714 | FZDD | 0 | 0 | 11 | 0 | - | FZDD | 1.00 | |
| 4: | 0 | 1 | 6 | 7 | .571 | ZD | 0 | 0 | 18 | 24 | - | ZD | 1.00 | |
| 5 | 0 | 0 | 11 | 11 | - | Z | 0 | 0 | 36 | 43 | - | Z | ||
| 6 | 0 | 0 | 4 | 4 | - | BD | 0 | 0 | 22 | 27 | - | BD | ||
| 7 | 0 | 0 | 2 | 2 | - | ZSD | - | - | - | - | - | - | ||
| 8 | 0 | 0 | 3 | 3 | - | BDD | 0 | 0 | 8 | 8 | - | BDD | ||
| 9 | 0 | 0 | 3 | 3 | - | ZDD | - | - | - | - | - | - | ||
| 10 | 0 | 0 | 4 | 4 | - | FZ | 0 | 0 | 21 | 27 | - | FZ | ||
| 11 | 0 | 0 | 1 | 1 | - | FZSD | - | - | - | - | - | - | ||
| 12 | 0 | 0 | 3 | 3 | - | FZD | 0 | 0 | 13 | 14 | - | FZD | ||
| 13 | 0 | 0 | 3 | 3 | - | FBDD | 0 | 0 | 3 | 2 | - | FBDD | ||
| 14 | 0 | 0 | 5 | 4 | - | MZ | 0 | 0 | 18 | 23 | - | MZ | ||
| 15 | 0 | 0 | 2 | 2 | - | MZSD | - | - | - | - | - | - | ||
| 16 | 0 | 0 | 4 | 4 | - | MZD | 0 | 0 | 13 | 14 | - | MZD | ||
| 17 | 0 | 0 | 1 | 2 | - | MBDD | 0 | 0 | 6 | 5 | - | MBDD | ||
| 18 | 0 | 0 | 2 | 3 | - | MZDD | - | - | - | - | - | - | ||
| 19 | - | - | - | - | - | - | 1 | 0 | 1 | 0 | - | FFBDD | ||
| 20 | - | - | - | - | - | - | 0 | 0 | 6 | 10 | - | BSD | ||
| 21 | - | - | - | - | - | - | 0 | 0 | 5 | 6 | - | FBSD | ||
| 22 | - | - | - | - | - | - | 0 | 0 | 6 | 7 | - | FFZ | ||
| 23 | - | - | - | - | - | - | 0 | 0 | 4 | 5 | - | FFBD | ||
| 24 | - | - | - | - | - | - | 0 | 0 | 3 | 2 | - | FFBSD | ||
| 25 | - | - | - | - | - | - | 0 | 0 | 2 | 3 | - | FFZD | ||
| 26 | - | - | - | - | - | - | 0 | 0 | 6 | 5 | - | MBSD | ||
| 27 | - | - | - | - | - | - | 0 | 0 | 10 | 11 | - | MFZ | ||
| 28 | - | - | - | - | - | - | 0 | 0 | 7 | 7 | - | MFBD | ||
| 29 | - | - | - | - | - | - | 0 | 0 | 4 | 3 | - | MFBSD | ||
| 30 | - | - | - | - | - | - | 0 | 0 | 7 | 9 | - | MFZD | ||
| 31 | - | - | - | - | - | - | 0 | 0 | 4 | 2 | - | MFBDD | ||
| 32 | - | - | - | - | - | - | 0 | 0 | 10 | 11 | - | MMZ | ||
| 33 | - | - | - | - | - | - | 0 | 0 | 5 | 4 | - | MMBD | ||
| 34 | - | - | - | - | - | - | 0 | 0 | 3 | 1 | - | MMBSD | ||
| 35 | - | - | - | - | - | - | 0 | 0 | 6 | 5 | - | MMZD | ||
| 36 | - | - | - | - | - | - | 0 | 0 | 2 | 1 | - | MMBDD | ||
| Key | |
| A | frequency of actual marriages with a given type of relative |
| S | frequency of simulated random marriages with a given type of relative |
| TA | total of actual relatives of this type |
| TS | total of simulated relatives of this type |
| p | probability (Fisher Exact) |
| Table 8: Test of Actual versus Simulated Marriage among Consanguineal Kin for Pul Eliya (conclusion: MBD is a preferred marriage) | |||||||||
| Type of | Actual Freq. | Simul Freq. | Total Actual | Total Simul | Fisher Exact | Blood Marriage | Viri-Sided? | ||
| Mar. | Type | P-graph notation | Actual | Simul | |||||
| 12: | 5 | 0 | 40 | 38 | .042 | MBD | GF=FG | yes | |
| 2: | 3 | 1 | 39 | 40 | .317 | FZD | GG=FF | yes | |
| 1: | 0 | 1 | 56 | 57 | .508 | FZ | GG=F | no | |
| 3: | 0 | 1 | 6 | 6 | .538 | FFFZDSD | GGGG=FGFF | no | |
| 4: | 1 | 0 | 3 | 1 | .800 | FFMZDSSD | GGGF=FGGFF | yes | |
| 5: | 0 | 1 | 5 | 3 | .444 | FFMBDSDD | GGGF=FFGFG | no | |
| 6: | 1 | 0 | 18 | 15 | .558 | FMBSD | GGF=FGG | yes | |
| 7: | 0 | 1 | 17 | 12 | .433 | FMBDD | GGF=FFG | no | |
| 8: | 2 | 1 | 18 | 12 | .661 | FMZDD | GGF=FFF | yes | |
| 9: | 0 | 1 | 9 | 5 | .399 | FMMBSSD | GGFF=FGGG | no | |
| 10: | 0 | 1 | 4 | 5 | .600 | FMMFZSSD | GGFFG=FGGF | yes | |
| 11: | 0 | 1 | 6 | 3 | .400 | FMMFZDSD | GGFFG=FGFF | yes | |
| 13: | 0 | 1 | 25 | 27 | .528 | MBSD | GF=FGG | yes | |
| 14: | 1 | 0 | 14 | 10 | .600 | MFZDD | GFG=FFF | yes | |
| 15: | 1 | 0 | 7 | 3 | .727 | MFFZDSSD | GFGG=FGGFF | yes | |
| 16: | 1 | 0 | 8 | 4 | .692 | MFFZDSD | GFGG=FGFF | yes | |
| 17: | 1 | 0 | 8 | 2 | .818 | MFMBDSSD | GFGF=FGGFG | yes | |
| 18: | 1 | 0 | 9 | 3 | .769 | MFMBDD | GFGF=FFG | yes | |
| 19: | 1 | 0 | 3 | 0 | 1.000 | MFMBDDDD | GFGF=FFFFG | yes | |
| 20: | 1 | 0 | 8 | 2 | .818 | MFMFZSSD | GFGFG=FGGF | yes | |
| 21: | 1 | 0 | 3 | 0 | 1.000 | MFMFZDDD | GFGFG=FFFF | yes | |
| 22: | 1 | 0 | 13 | 8 | .636 | MMZSSD | GFF=FGGF | yes | |
| 23: | 1 | 0 | 15 | 13 | .551 | MMBDD | GFF=FFG | yes | |
| 24: | 0 | 1 | 11 | 5 | .352 | MMZSDD | GFF=FFGF | no | |
| 25: | 0 | 1 | 11 | 5 | .352 | MMBDDD | GFF=FFFG | no | |
| 26: | 1 | 0 | 11 | 4 | .749 | MMZDDD | GFF=FFFF | yes | |
| Table 9: non-MBD marriages, Correlating Actual versus Simulated with Dravidian Viri-Sided/Unsided Marriage (p = 0.0004; p = 0.000004 using the binomial test of 50%:50% expected) | ||
| Viri-Sided | Unsided | |
| Actual | 18 | 0 |
| Simulated | 5 | 7 |
| Table 10: non-MBD generationally "skewed" marriages, Correlating Actual versus Simulated with Dravidian Uxori-Sided/Unsided Marriage (p=.02; >p=.002 using the binomial test of 50%:50% expected) | ||
| Uxori-Sided | Unsided | |
| Actual | 0 | 9 |
| Simulated | 4 | 3 |
| Table 11: Test of Sidedness for Pul Eliya (Programs: PGRAPH and Par-Side) Number of Elementary Cycles: 4 | ||
| A. Viri-sidedness | Actual | Expected |
| Balanced Cycles (Even length) | 25 | 17.5 |
| Unbalanced Cycles (Odd Length) | 10 | 17.5 |
| p = 0.008 | ||
| B. Amblilateral-sidedness (women adjusted by inheritance rules) | Actual | Expected |
| Balanced Cycles (Even length) | 35 | 17.5 |
| Unbalanced Cycles (Odd Length) | 0 | 17.5 |
| p = 0.00000000003 | ||
| Table 12: Test of Sidedness for Javanese Muslim Elites (Programs: PGRAPH and Par-Side) Number of Elementary Cycles: 4 | ||
| A. Viri-sidedness | Actual | Expected |
| Balanced Cycles (Even length) | 1 | 2 |
| Unbalanced Cycles (Odd Length) | 3 | 2 |
| P = 0.94 | ||
| B. Uxori-sidedness | Actual | Expected |
| Balanced Cycles (Even length) | 3 | 2 |
| Unbalanced Cycles (Odd Length) | 1 | 2 |
| P = 0.31 | ||
| Table 13: Comparison of Relinking Frequencies for Actual and Simulated Data (*=greater than chance), from Brudner and White (1997) | ||||||||||||
| Magnitude of Structural Endogamy with ancestors back 1, 2, ..., g generations | ||||||||||||
| Back: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Starting from: | ||||||||||||
| Present generation | ||||||||||||
| Actual | 8* | 16* | 70* | 179 | 257 | 318 | 349 | 363 | 376 | 390 | 399 | 405 |
| Simulated | 0 | 0 | 32 | 183 | 273 | 335 | 365 | 382 | 388 | 397 | 397 | 403 |
| Back one | ||||||||||||
| Actual | 8* | 58* | 168 | 246 | 308 | 339 | 353 | 366 | 380 | 389 | 395 | |
| Simulated | 0 | 18 | 168 | 255 | 320 | 347 | 366 | 372 | 381 | 381 | 387 | |
| Back two | ||||||||||||
| Actual | 26* | 115* | 178 | 243 | 278 | 292 | 305 | 319 | 328 | 334 | ||
| Simulated | 0 | 98 | 194 | 262 | 291 | 310 | 316 | 325 | 325 | 331 | ||
| Table 14: Test of Actual versus Simulated Marriage among Consanguineal Kin for Austrian Village (conclusion: maternal-side blood marriages are avoided, paternal side not) | |||||||
| A | S | TA | TS | P= | |||
| Type of | Actual Freq. | Simul Freq. | Total Actual | Total Simul | Fisher Exact | Blood Marriage | |
| Mar | type | degree | |||||
| 7: | 1 | 0 | 32 | 34 | .492 | FFMBDD | 2nd + |
| 1: | 1 | 0 | 165 | 144 | .535 | FBD | 1st |
| 14: | 1 | 0 | 34 | 33 | .514 | FMMBSD | 2nd - |
| 8: | 1 | 0 | 31 | 28 | .533 | FFMZDD | 2nd + |
| 17: | 1 | 0 | 12 | 11 | .541 | FMMMBSSD___ | 3rd + median |
| 13: | 1 | 0 | 6 | 5 | .583 | FMFMZSDD | 3rd ++ |
| 16: | 1 | 0 | 4 | 3 | .625 | FMMFZDDSD | 4th |
| 23: | 1 | 0 | 3 | 2 | .666 | MFMMBSSDD | |
| 24: | 1 | 0 | 54 | 22 | .714 | MMBSD | |
| 26: | 1 | 0 | 8 | 3 | .750 | MMMBSDD | |
| Mo's side | Fa's side | ||
| <.55 (median) | 0 | 5 | |
| >.55 | 3 | 2 | p=.08 |
| 25: | 0 | 1 | 23 | 5 | .206 | MMFBSSD | 3rd |
| 21: | 0 | 1 | 10 | 3 | .285 | MFMFBSSD | 3rd + |
| 20: | 0 | 1 | 16 | 6 | .304 | MFMBDSD | 3rd |
| 19: | 0 | 1 | 6 | 2 | .333 | MFFFBDSD | 3rd + |
| 5: | 0 | 1 | 20 | 13 | .411 | FFFZSDD | 3rd |
| 18: | 0 | 1 | 7 | 4 | .416 | MFBDDDD | 2nd -- |
| 9: | 0 | 1 | 5 | 3 | .444 | FFMFBDDSD | 4th |
| 15: | 0 | 1 | 21 | 16 | .447 | FMMZSSD____ | 3rd median |
| 11: | 0 | 1 | 46 | 37 | .452 | FMBSSD | |
| 2: | 0 | 1 | 105 | 91 | .467 | FBSD | |
| 6: | 0 | 1 | 17 | 16 | .499 | FFFZDDD | |
| 10: | 0 | 1 | 4 | 3 | .500 | FFMFBDSDD | |
| 12: | 0 | 1 | 10 | 10 | .523 | FMFFZSSD | |
| 3: | 0 | 1 | 16 | 21 | .578 | FBSDDD | |
| 4: | 0 | 1 | 10 | 13 | .583 | FFFBSSDD | |
| 22: | 0 | 1 | 1 | 1 | .666 | MFMFBDSDD |
| Mo's side | Fa's side | ||
| <.42 | 5 | 3 | |
| >.42 | 1 | 7 | P = 0.06 |
| Mo's side | Fa's side | ||
| Actual | 0 | 5 | |
| Simulated | 5 | 3 | p=.04 |
| key: | |
| A = | frequency of actual marriages with a given type of relative |
| S = | frequency of simulated random marriages with a given type of relative |
| TA = | total of actual relatives of this type |
| TS = | total of simulated relatives of this type |
| p = | probability (Fisher Exact) |
| Table 15: Comparison of Magnitude of Attribute versus Structural Endogamy as Predictors | ||
| A. attribute endogamy: language | ||
| marriage | Wi bilingual | monolingual |
| Hu bilingual | 96 | 16 |
| monolingual | 7 | 15 |
| r=.47 | ||
| B. attribute endogamy: occupation | ||
| marriage | Wi's fa: farmer | non-farmer |
| Hu's fa: farmer | 80 | 7 |
| non-farmer | 15 | 34 |
| r=.64 | ||
| C. structural endogamy | ||
| Marriage | block members | non-block |
| Heirs | 173 | 117 (adjusted est.40) |
| Residents | 25 | 281 |
| r=.55, adj. r=.74 | ||
| 3-way tests of difference | A&B p=.46 |
| B&C adjusted, p=.000006 (greater correlation for structural endogamy) | |
| A&C adjusted, p=.000007 (greater correlation for structural endogamy) |
Acknowledgements
Notes2Paths that are not contained in circuits are trees which can always - trivially - be mapped onto a bipartite graph by assigning a simple alternation of connected nodes to supersets.
3There is a consequent dearth of current applications of his programs to the issues of marriage rules and strategies, partly caused by a failure to supply a personal-computer version of the software and perhaps also through lack of motivation given Hammel's (1976b) finding.
4This approach is being pursued through a working group at the Santa Fe Institute.
5The more standard approach of Monte Carlo estimation is to run many simulations, build a statistical distribution empirically, and then locate where the observed sample is located in this distribution to give a probability estimate. While studies such as that of White et al (1999) used this approach, it is computationally cumbersome and, as in the present case, unnecessary. The computer programs used here can, of course, be used to verify that the present approach converges with Monte Carlo results, but that will not be undertaken here.
6If Leach was later stung by Hammel's (1976b) critique, with its mocking title, of the Leach 1951 arguments about MBD as a strategic alliance among the Kachin, structurally controlled simulation using permutation tests provides a means of refuting Hammel's argument concerning particular cases of MBD marriage.
Appendix: A List of Relevant Computer Programs
BATAGELJ, Vladimir and Andrej Mrvar. 1997. Networks / Pajek: Program for Large Networks Analysis. University of Ljubljana, Slovenia. http://vlado.fmf.uni-lj.si/pub/networks/pajek/.
BRUDNER, Lilyan A. 1969. The Ethnic Component of Social Transactions. Ph.D. Dissertation: University of California, Berkeley.
BRUDNER, Lilyan A. and Douglas R. White. 1997. Class, Property and Structural Endogamy: Visualizing Networked Histories. Theory and Society 25:161-208.
GIBBONS, Alan. 1985. Algorithmic graph theory. Cambridge: University Press.
HAMMEL, Eugene.1976a. The SOCSIM demographic-sociological microsimulation program: operating manual. Berkeley: Institute of International Studies, University of California.
HAMMEL, Eugene. 1976b. The Matrilateral Implications of Structural Cross-Cousin Marriage. pp. 145-168, in, Ezra B. W. Zubrow, editor, Demographic Anthropology: A Quantitative Approach. Albuquerque: School of American Research, University of New Mexico Press.
HOUSEMAN, Michael. 1997. Marriage networks among Australian Aboriginal populations. Australian Aboriginal Studies 2: 2-23.
HOUSEMAN, Michael, and Douglas R. White. 1996. Structures réticulaires de la pratique matrimoniale. L'Homme 139:59-85.
HOUSEMAN, Michael, and Douglas R. White. 1998a. Ambilateral Sidedness among the Sinhalese: Marriage Networks and Property Flows in Pul Eliya, pp. 59-89 in Thomas Schweizer and Douglas R. White, eds.Kinship, Networks and Exchange. Cambridge University Press.
HOUSEMAN, Michael, and Douglas R. White. 1998b. "Taking Sides: Marriage Networks and Dravidian Kinship in Lowland South America," pp. 214-243 in, Maurice Godelier and Thomas Trautmann, eds., Transformations of Kinship. Washington, D.C.: Smithsonian Institution Press.
JOLA, Tina, Yvonne Verdier and Françoise Zonabend. 1970. Parler famille. L'homme 10/3:5-26.
LANG, Hartmut. 1995. Demographic Micro-Simulation. Paper presented at the Nanterre conference of the Working Group on Kinship and Computing, September, 1995.
LEACH, Edmund R. 1951. The Structural Implications of Matrilateral Cross-Cousin Marriage. Journal of the Royal Anthropological Institute 81:23-55.
LEACH, E. R. 1961 [1968]. Pul Eliya: A Village in Ceylon. Cambridge: University Press.
LÉVI-STRAUSS, Claude. 1966. The Future of Kinship Studies. Proceedings of the Royal Anthropological Institute for 1965. pp. 13-22.
REED, R. 1974. "Relative Age and Asymmetrical Cross-Cousin Marriage in a South Indian Caste," pps. 257 - 274, in, Paul Ballonoff, ed., Genealogical Mathematics. Paris: Maison des sciences de l’homme.
RICHARD, Philippe. 1993. "Étude des renchaînements d'alliance," Mathématique, Informatique et Science humaines 123:5-35.
ROMNEY, A. Kimball 1971. Measuring of Endogamy. pp. 191-213, in, Paul Kay (ed.)Explorations in Mathematical Anthropology. Cambridge: MIT Press.
SCHWEIZER, Thomas. 1989. Reisanbau in einem javanischen Dorf: Eine Fallstudie zu Theorie und Methodik der Wirtschaftsethnologie. Cologne: Böhlau Verlag.
SEGALEN, Martine. 1985. Quinze générations des Bas-Bretons. Parenté et société dans le pays bigouden sud. 1720-1980. Paris: Presses Universitaires Français.
WHITE, Douglas R. 1973. Mathematical Anthropology, pp. 369-446, in, John J. Honigmann, editor, Handbook of Social and Cultural Anthropology. Chicago: Rand-McNally and Co.
WHITE, Douglas R. 1994. Fisher-B: A Program for Exact Significance Tests for Two and Three-Way Interaction Effects. World Cultures 8(2):40-43.
WHITE, Douglas R. 1997. Structural Endogamy and the graphe de parenté. Mathématique, Informatique et Sciences Humaines.137:107-125.
WHITE, D. R., Michael Schnegg, Lilyan A. Brudner, and Hugo G. Nutini. 1999. "Multiple Connectivity and its Boundaries of Integration: A Community Study." University of California - Irvine. Spanish translation (in press): Conectividad Múltiple y sus Fronteras de Integración: Parentesco y Compadrazgo en Tlaxcala Rural. In, Jorge Gil and Samuel Schmidt, eds., Redes Sociales: Teoría y Aplicaciones. México, DF: UNAM Press.
WHITE, Douglas R., and Michael Houseman. book ms. Balance in Kinship Networks: Cognatic Dual Organization.
WHITE, Douglas R., and Paul Jorion. 1992. Representing and Analyzing Kinship: A Network Approach. Current Anthropology 33:454-462.
WHITE, Douglas R., and Paul Jorion. 1996. Kinship Networks and Discrete Structural Analysis: Formal Concepts and Applications. Social Networks 18:267-314.
WHITE, Douglas R., Robert Pesner, and Karl Reitz. 1983. An Exact Test for 3-Way Interactions. Behavior Science Research 17:103-122.
WHITE, Douglas R., and Thomas Schweizer. 1998. Kinship, Property Transmission, and Stratification in Rural Java, pp. 36-58 in Kinship, Networks and Exchange, edited by Thomas Schweizer and Douglas R. White. Cambridge University Press. In Press.
WHITE, Douglas R., and Patricia Skyhorse. 1996. PGRAPH User's Manual. University of California Irvine.
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, 1999