
Federico Cecconi and Domenico Parisi (1998)
Journal of Artificial Societies and Social Simulation vol. 1, no. 2, <https://www.jasss.org/1/2/1.html>
To cite articles published in the Journal of Artificial Societies and Social Simulation, please reference the above information and include paragraph numbers if necessary
Received: 24-Oct-1997 Accepted: 12/2/98 Published: 31/3/98
 Abstract
Abstract
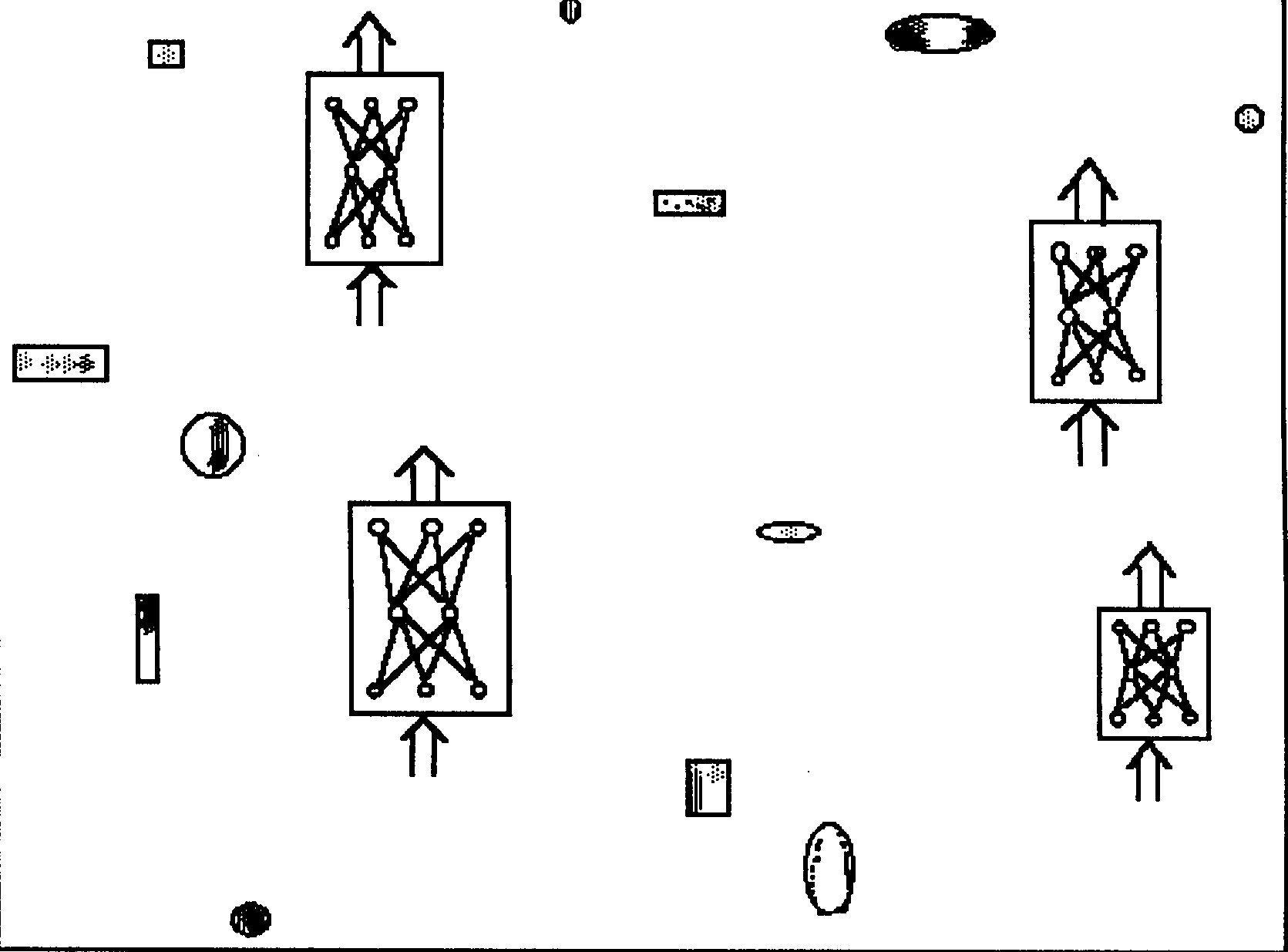
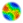
Figure 1. A group of individuals adopting an individual survival strategy.
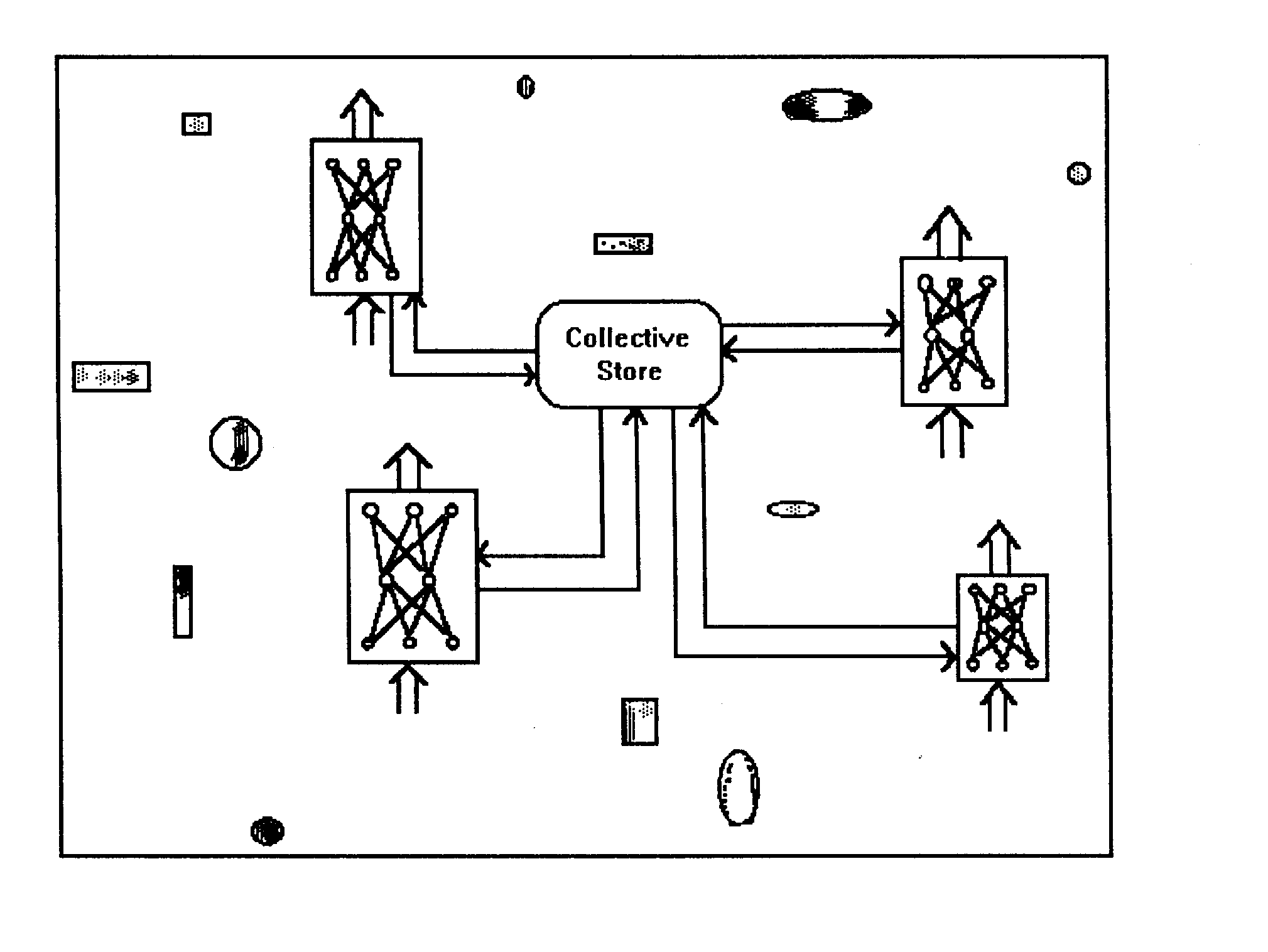
Figure 2. A group adopting a Central Store strategy. The individuals contribute (some of) the food they are able to capture in the environment to a Central Store and the food contained in the Central Store is redistributed to the entire group.
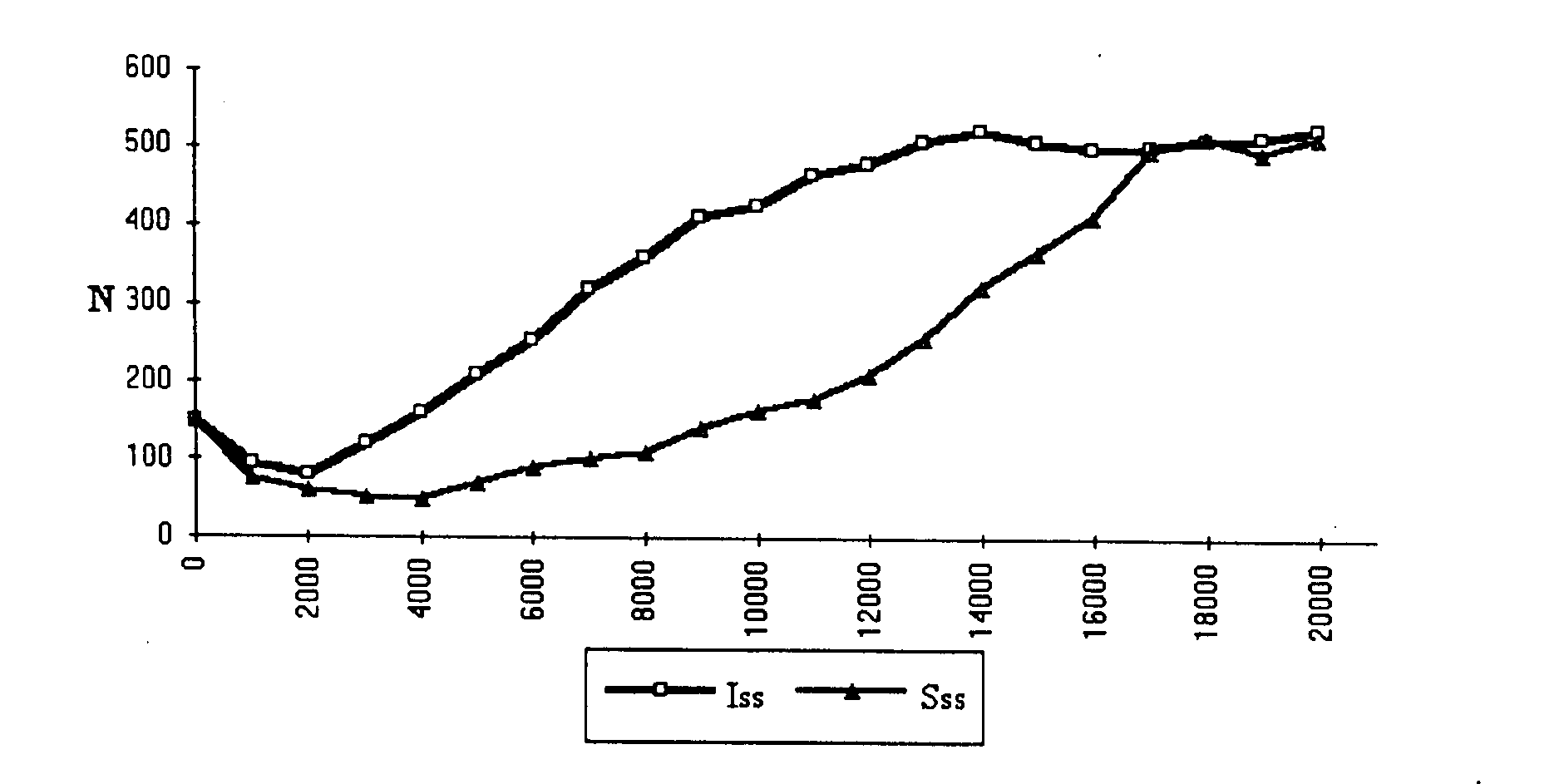
Figure 3. Evolutionary change in group size in ISS and CS groups across 20,000 cycles.
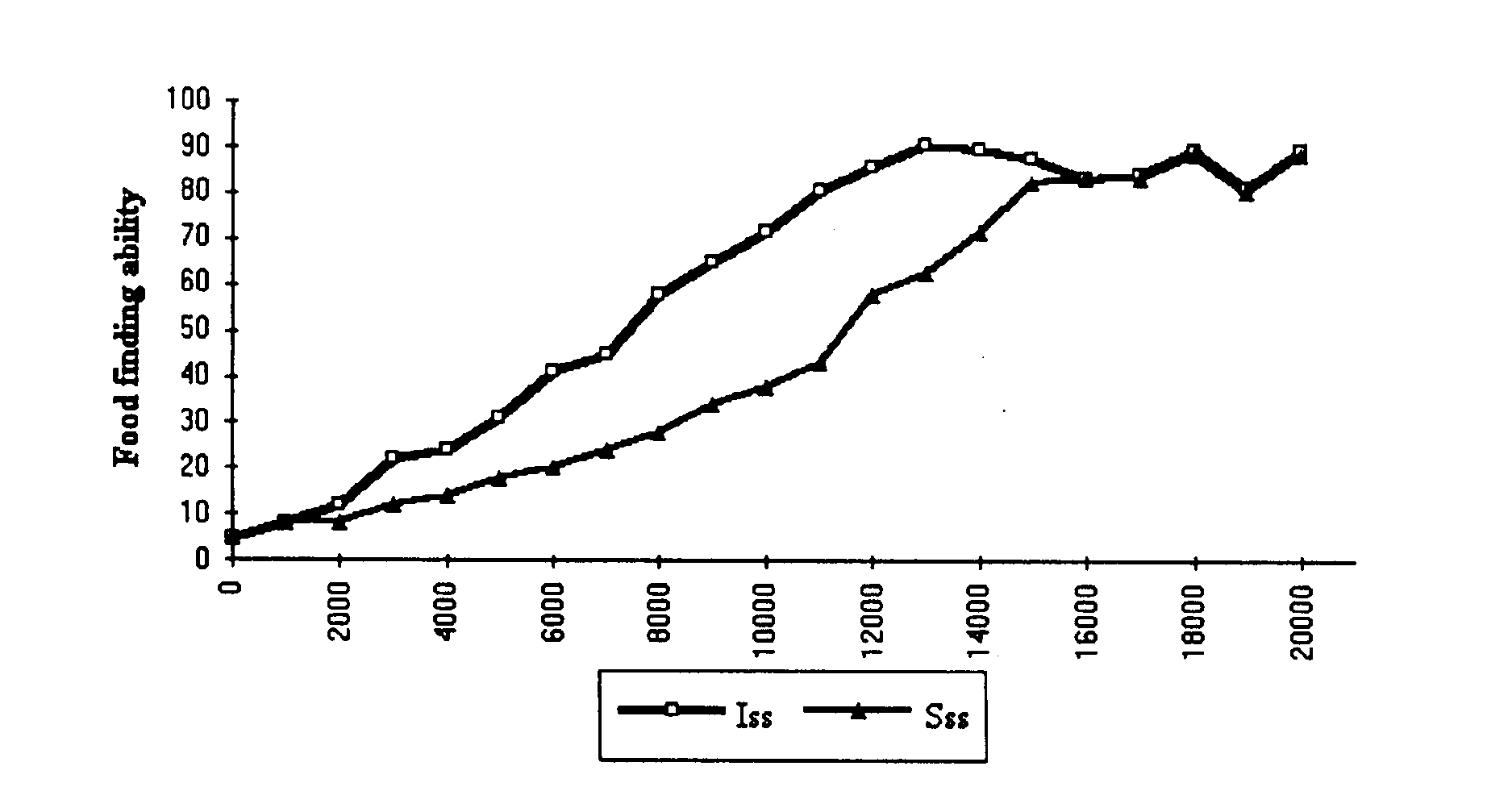
Figure 4. Evolutionary change in food capturing ability in ISS and CS groups. The food capturing ability of individuals belonging to the two groups is tested in identical 'laboratory' conditions
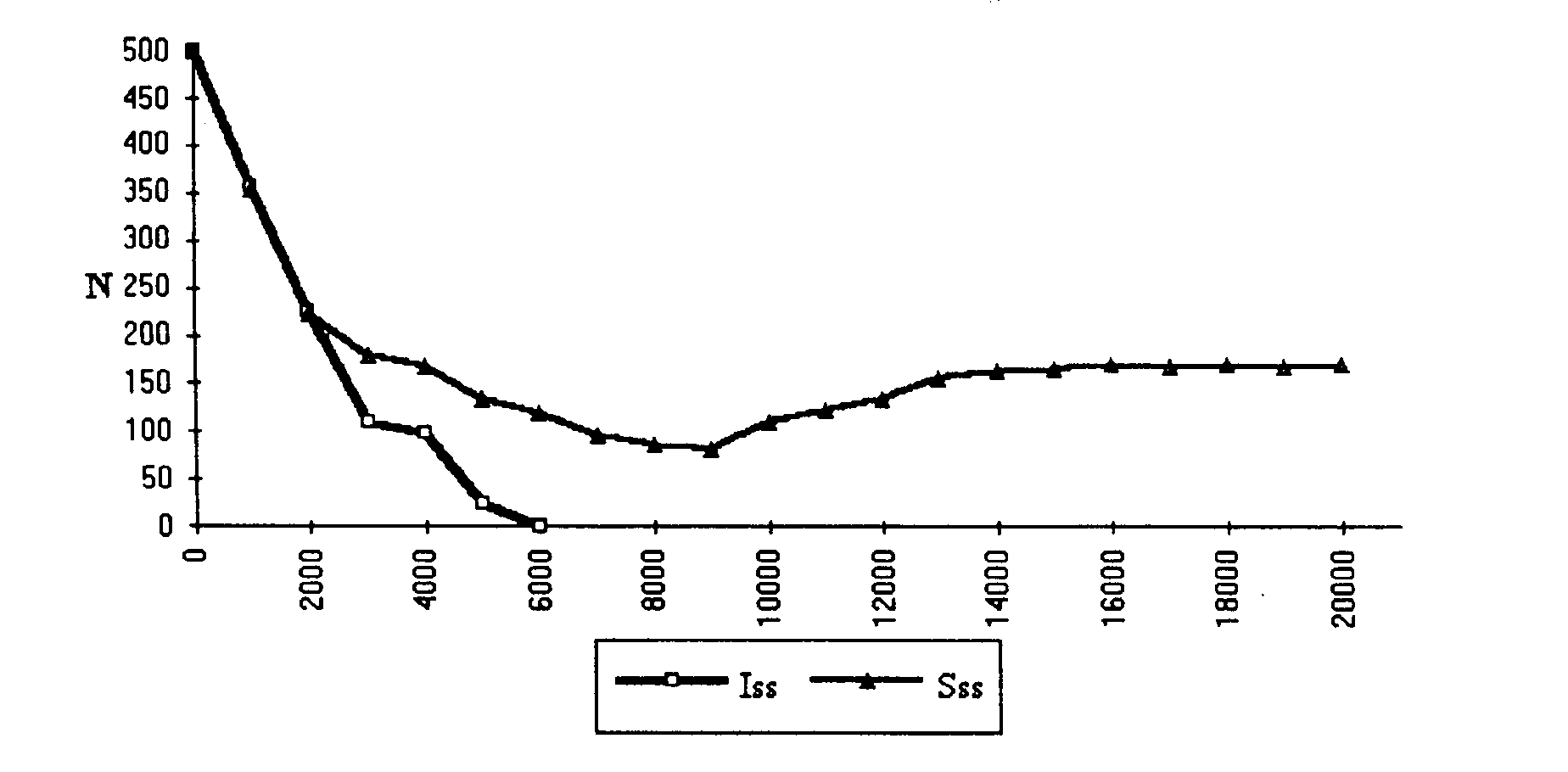
Figure 5. Change in group size in ISS and CS groups after a worsening of environmental conditions. The ISS groups all become extinct after 6,000 cycles (average of 25 runs of the simulation) while all the CS groups succeed in avoiding extinction and in reaching a new stable state corresponding to the carrying capacity of the new environment
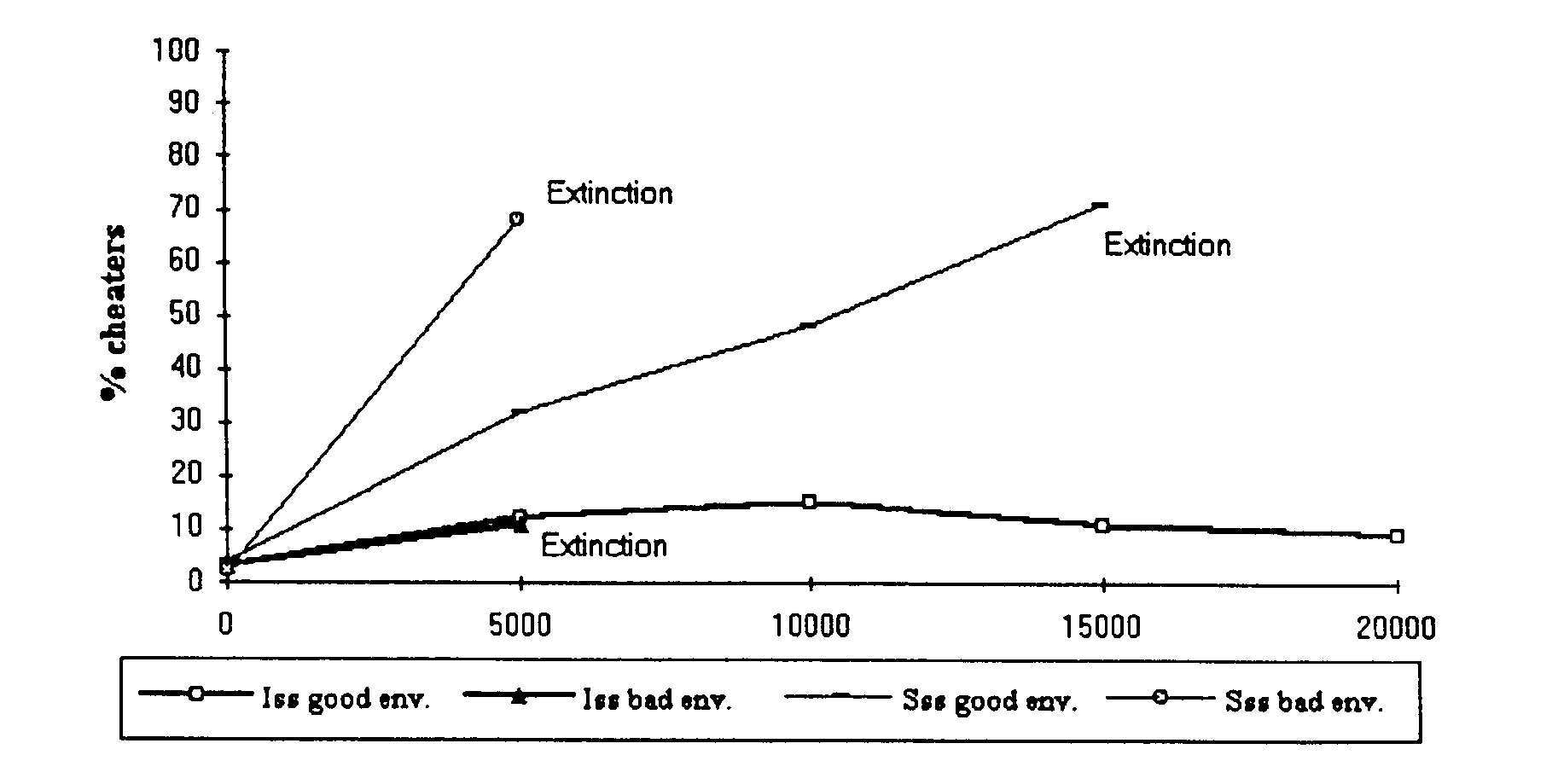
Figure 6. Percentage of 'do nothing' actions in each cycle across 20,000 cycles of evolution for ISS and CS groups living in either favourable or unfavourable environments. All groups become extinct except the ISS groups living in the favourable environment, which succeed in keeping the percentage of 'do nothing' actions at very low levels.
CECCONI, F. and PARISI, D. Learning during reproductive immaturity in evolving populations of neural networks. Institute of Psychology, CNR, 1994.
COLSON, E. In good years and bad: food strategies of self-reliant societies. Journal of Anthropological Research, 1980, 35, 1-29.
EARLE, T.K. How Chiefs Come to Power. The Political Economy in Prehistory. Stanford, Stanford University Press, 1997.
HAMILTON, W.D. The genetical evolution of social behavior. Journal of Theoretical Biology, 1964, 7, 1-16.
INGOLD, T. The significance of storage in hunting societies. Man, 1983, 18, 553-571.
PARISI, D. What to do with a surplus. In R. Conte, P. Terna, & R. Hegselmann (eds.) Simulating Social Phenomena. Berlin, Springer, 1997.
PEDONE, R. and PARISI, D. In what kinds of social groups can "altruistic" behaviors evolve? In R. Conte, P. Terna, & R. Hegselmann (eds.) Simulating Social Phenomena. Berlin, Springer, 1997.
RAUP, D.M. Extinction. New York, Norton, 1991.
STEWARD, J. Evolutionary Transitions and Artificial Life. Artificial Life, 1997, 3, 101-120
TESTART, A. The significance of food storage among hunter-gatherers: residence patterns, population densities, and social inequalities. Current Anthropology, 1982, 25, 523-537.
TRIVERS, R. Social Evolution. Menlo Park, CA, Benjamin/Cummings, 1985.
WILSON, D.S. and SOBER, E. Reintroducing group selection to the human behavioral sciences. Behavioral and Brain Science, 1994, 17, 585-608.
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, 1998