
Dwight W. Read (1998) 'Kinship based demographic simulation of societal processes'
Journal of Artificial Societies and Social Simulation vol. 1, no. 1, <https://www.jasss.org/1/1/1.html>
To cite articles published in the Journal of Artificial Societies and Social Simulation, please reference the above information and include paragraph numbers if necessary
Received: 29-Oct-1997 Accepted: 12-Dec-1997 Published: 3-1-1998
 Abstract
Abstract
| dP(t)/dt = rP(t)(1 - P(t)/K) | (1) |
where P(t) is the population size at time t, dP(t)/dt is the (instantaneous) rate of growth at time t, r is the net growth rate per female and K is usually interpreted as the carrying capacity. (Or, in words, Equation 1 asserts that the rate of change of population size is proportional to the current population size, but with the proportionality -- the term r(1-P(t)/K) -- decreasing as the population size increases.) The vector, P = <P> is one dimensional; the parameters are r and K, and Equation (1) makes up the set, E, of structural equations and defines the behavior of the system. Equation 1 and variants on it have been widely used for modeling growth in populations. In the deterministic, non-stochastic situation, the system is driven asymptotically to a stable equilibrium value, Pe = K, for r > 0. For groups such as the !Kung San this conclusion is consistent with the observation that hunting and gathering societies necessarily have bounded populations over long time scales (tens to hundreds of generations) by virtue of being constrained by a limited resource base. Equation 1 does not, however, provide a very good model of the process by which population dynamics are played out for a group such as the !Kung San.
Modeling Strategy: Minimum Specification
Assumption 1: !Kung San women have a desire for as many children as possible
and
Assumption 2: !Kung San women are concerned for, and make decisions to promote, the well-being of their family,
where family well-being includes, but is not limited to, ability of a woman to adequately nurse a newborn and to carry infants with her when foraging for resources. These two assumptions are based on comments elicited from !Kung San women by Lorna Marshall as discussed above.
Simulation Model
Fertility Models
| r = r0(1 - P/K) | (2) |
where K is a parameter that specifies the equilibrium population size, r0 is the intrinsic fertility rate (r0 = 8 births per female per reproductive period in all simulations), r is the current, realized fertility rate (note that in equation (2) r is the fertility rate whereas in equation (1) r is the net growth rate) and P is the number of adult women. The model separates the net growth rate of equation (1) into two parts, the realized fertility rate which is assumed to be density dependent and a fixed, age dependent mortality rate. Consequently, the parameter, K, in equation (2) is an overestimate of the stabilized population size since equation (2) does not include mortality rates. A fixed, age-specific mortality schedule is used in this and all other simulations.
| E = n*Wt + P/K | (3) |
where E is the total energy expenditure for a woman, n is the number of children below the infant age, IA, Wt is a weighting factor that represents the energy expenditure per infant, and K is a constant that translates population size into energy cost equivalents per women per infant (and represents in summary fashion her total cost in doing all activities other than caring of infants).
| r = 0 if E >= T |
| r = r0if E < T | (4) |
| IA= 4 *(P/K) | (5) |
Equation 5 relates a woman's current willingness to have an offspring to the time/energy she must expend on obtaining resources. The energy expenditure per woman is computed in the same manner as for Model 2:
| E = n*Wt + P/K | (6) |
and equation (4) is used for assigning the current fertility rate of a woman. This model captures the notion that a choice to have another child is affected by both (1) the current amount of time/energy spent on obtaining resources (which affects the number of years she will nurse, hence the age for an infant) and (2) her cost of having currently having n infants.
Population Growth And Birth Spacing: Results
Birth Spacing
|
TABLE 1 |
|||
| AVERAGE BIRTH SPACING | |||
| Model | Mean (x) | Standard Deviation (s) | Number of Females |
| Model 1 | 3.70 | 1.84 | 306 |
| Model 2c | 3.47 | 1.46 | 249 |
| Model 3 | 3.90 | 1.31 | 201 |
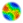
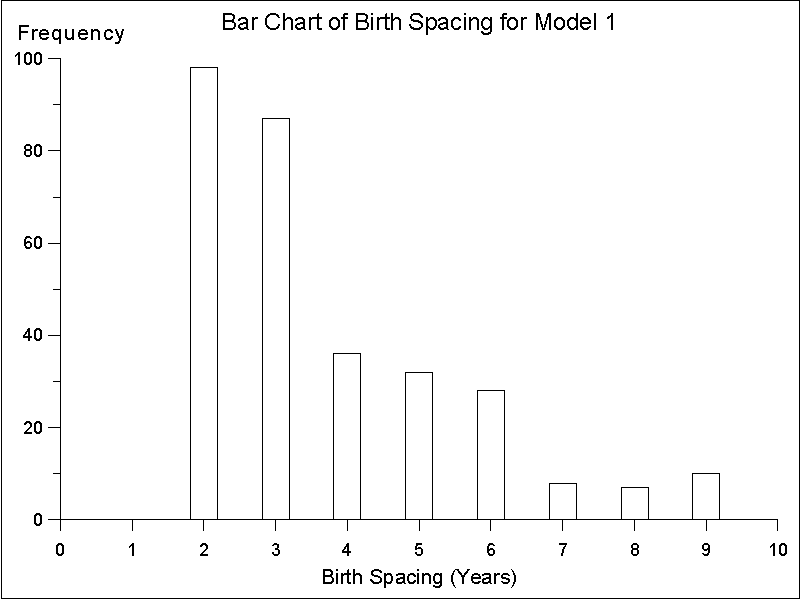
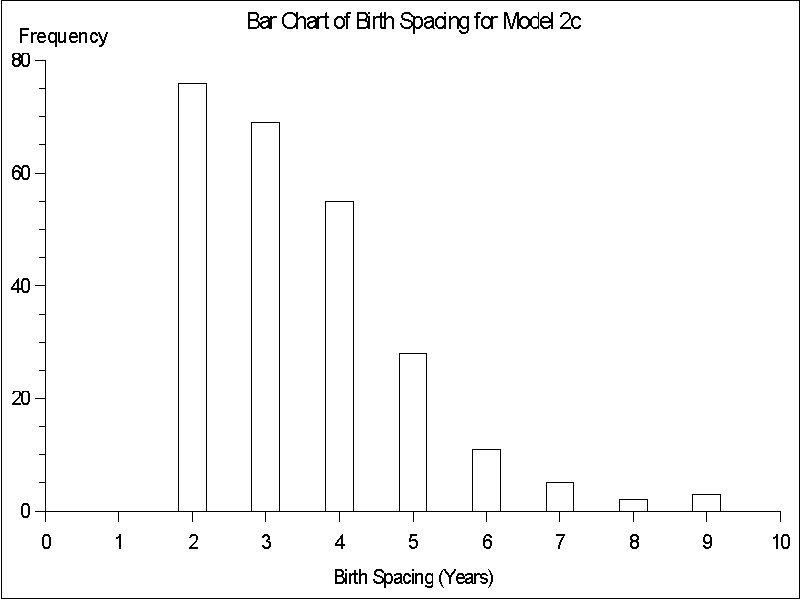
44.1 month average spacing (n = 55) observed for the more nomadic !Kung San women (Lee 1980: 336). Differences among the models arise, however, in the frequency distribution of birth spacing for each model. For Model 1, spacings of 2 or 3 years are most common, with longer spacing occurring substantially less often (see Figure 8). In Model 2, frequency of spacing drops off more smoothly, with spacing of 2 - 4 years the most common values, reflecting the fact that when fertility changes at the threshold value all women are affected equally regardless of their current family structure (see Figure 9). In Model 3 the modal value is 3 year spacing with spacing of 4 years also common. Spacing fewer than 3 years is rare (see Figure 10). Thus, although the average birth spacing is comparable across the three models (which primarily reflects the fact that all three models arrive at stabilized population values), only Model 3 leads to a birth spacing frequency distribution that approximates the actual birth spacing by !Kung San women. Common values for birth spacing are 3 and 4 years and seldom is less than 3 years (Howell 1976: 145).
Marriage Rules
!Kung San Kinship Terminology
Simulation Incest Rule: Marriage between ego and alter will be incestuous if alter is in any of the kinship lists of ego, and vice-versa.
Kin Terms: Implementation
!Kung San Naming System: Implementation
Camp Membership
Rule: Stay in her camp if: her camp size < 30 or his natal camp size > 30, but relocate to his camp if her camp size > 30 and his natal camp size is < 30.
The value of 30 is used as camps tend to average about 30 persons.
Results: Implications Of Incest Rules
|
TABLE 2 |
||
| Incest Type | Marriages in Camp/Total Marriages | Total Population |
| Model 3a (!gun!a Incest) | 2/91 = 0.02 | 270 |
| Model 3b (Cousin Incest) | 16/67 = 0.24 | 243 |
| Model 3c (Parental Incest) | 22/74 = 0.30 | 281 |
in the least restrictive case, Model 3c, where only parent, parents' siblings, siblings and children are incestuous, about 1/3 of the marriages are within camps. Adding cousin incest (1st and 2nd cousins) produces Model 3b and the added incest has a relatively minor additional effect on camp exogamy. With !gun!a incest included (Model 3a), that is incest that includes close relatives for whom a female uses the term !gun!a, there is virtually complete camp exogamy (and in other runs there were either 0 or at most 1 endogamous marriages). Evidently a cultural rule of making close relatives for whom the term !gun!a is used incestuous suffices to create de facto camp exogamy. That there are pragmatic reasons for making camp exogamy efficient with respect to the distribution of persons over resources via camp membership suggests that the cultural rule may be the result of interplay between the kinship system as a cultural phenomenon, on the one hand, and the extent to which redistribution of persons in camps is aided or hindered by culturally specified incest rules.
Conclusion
1Similar comments about the difficulty of providing enough food for a family have been elicited from other hunter/gatherer groups. For example, with reference to groups in South Australia, Tiechelmann writes: "A mother who had already killed two of her children said -- "how could I accompany my husband, and how could I supply food enough were I to bring up so many children?" (Tiechelmann 1841: 13) quoted in (Hayden 1972: 213).
2The relationship between months of breast feeding and months of amenorrhea is well-illustrated by worldwide data on women from 21 countries provided by (Bongaarts and Potter 1983). Their data are well fit by a second degree polynomial that implies an almost 1-1 equivalence between months of breast feeding and months of amenorrhea when breast feeding extends beyond about 30 months.
3Intensification of foraging and hunting can lead to degradation of resource availability. Habitat productivity, except for a few examples such as the use of fire to create more open country habitat, is not augmented via intensification of foraging and hunting procedures as can occur with agricultural production and intensification.
4It should be noted that, as demonstrated in the simulations, an average spacing of 4 years will arise when a population with a mortality schedule and actual reproductive period for women comparable to that of the !Kung San has a zero net growth rate, regardless of the factors affecting the spacing of offspring. This follows from the fact that the average spacing of offspring that must occur in order to maintain a zero net growth rate is determined by the mortality schedule and the length of the actual reproductive period. Thus the average birth spacing can be treated as responding to adaptive pressures only if it is assumed there are no factors driving the population size to an equilibrium value.
5While Howell (1976: 147) has argued that !Kung San women do not control their fertility artificially through means such as herbal teas, this does not rule out the possibility that !Kung San women can affect time of next pregnancy through a decision to continue or terminate breast-feeding. The strong evidence for the role of breast-feeding, particularly as practiced by !Kung San women, in post-partum amenorrhea along with the keen awareness of hunting and gathering groups about the biological world make it unlikely that they are unaware of the role of breast-feeding in producing temporary fecundity reduction. Such awareness may be mediated culturally in terms of how it is expressed; e.g., the quote presented above regarding the need to nurse for 3 to 4 years to ensure strong legs
6Gage et al. noted correctly that lactational amenorrhea, by itself, does not regulate population growth. They further argued that no link between lactational amenorrhea and a nonconservative factor (i.e., an extrinsic factor whose quantity varies with population density) had been established by those that proposed not only the existence of lactational amenorrhea but lactational amenorrhea as a causative mechanism in population regulation.
7In reality, other kin relations such as sibling relations are also used to justify camp membership.
8The number of camps relates to the number of permanent waterholes.
BALIKCI, Asen 1970 The Netsilik Eskimo. Gardern City, NY: The Natural History Press.
BLURTON Jones, Nicholas 1986 Bushman Birth Spacing: A Test for Optimal Interbirth Intervals. Ethology and Sociobiology 7:91-105.
BONGAARTS, J., and R. G. Potter 1983 Fertility, Biology and Behavior. New York: Academic Press.
ELLISON, P. T. 1994 Advance in Human Reproductive Ecology. In Annual Review of Anthropology, Vol. 23. William H. Durham, ed. Pp. 255-275. Palo Alto, California: Annual Reviews Inc.
GAGE, Timothy B., John M. McCullough, Charles A. Weitz, James S. Dutt, and Andrew Abelson 1989 Demographic Studies and Human Population Biology. In Human Population Biology: A Transdisciplinary Science. Michael A. Litte and Jere D. Haas, eds. Pp. 45-65. Oxford, England: Oxford University Press.
HAYDEN, Brian 1972 Population Control Among Hunter/Gatherers. World Archaeology 4:205-221.
HENRY, L.1961 Some Data on Natural Fertility. Eugenics Quarterly 8:81-91.
HOWELL, Nancy 1976 The Population of the Dobe Area !Kung. In Kalahari Hunter-Gatherers. Richard B. Lee and Irven DeVore, eds. Pp. 137-151. Cambridge, Massachusetts: Harvard University Press.
HOWELL, Nancy, and Victor Lehotay 1978 AMBUSH: A Computer Program for Stochastic Microsimulation of Small Human Populations. American Anthropologist 80(4):905-922.
JOHNSON, Gregory Alan 1983 Decision-making organization and pastoral nomad campsize. Human Ecology 11(2): 175-199.
KEESING, Roger 1973 Theories of Culture. Annual Review of Anthropology 3:73-97.
KONNER, M., and C. Worthman 1980 Nursing Frequency, Gonadal Function, and Birth Spacing Among !Kung Hunter-Gatherers. Science 207:788-791.
LEE, Richard B. 1976 !Kung Spatial Organization: An Ecological and Historical Perspective. In Kalahari Hunter-Gatherers. Richard B. Lee and Irven DeVore, eds. Pp. 73-97. Cambridge, Massachusetts: Harvard University Press.
LEE, Richard B. 1980 Lactation, Ovulation, Infanticide, and Women's Work: Study of Hunter-Gatherer Population Regulation. In Biosocial Mechanisms of Population Regulation. M.N. Cohen, R.S. Malpass and H.G. Klein, eds. Pp. 321-348. New Haven: Yale University Press.
LESLIE, Paul W., and Timothy B. Gage 1989 Demography and Human Population Biology: Problems and Rogress. In Human Population Biology: A Transdisciplinary Science. Michael A. Litte and Jere D. Haas, eds. Pp. 15-44. Oxford, England: Oxford University Press.
MARSHALL, Lorna 1976 The !Kung of Nyae Nyae. Cambridge, MA: Harvard University Press.
READ, Dwight W. 1974 Kinship Algebra: A Mathematical Study of Kinship Structure. In Genealogical Mathematics. Paul Ballonoff, ed. Pp. 135-161. Mouton Press.
READ, Dwight W. 1984 An Algebraic Account of the American Kinship Terminology. Current Anthropology 25:417-440.
READ, Dwight W. 1986 Mathematical Schemata and Archaeological Phenomena: Substantive Representation or Trivial Formalism? Science and Archaeology 38:16-23.
READ, Dwight W., and Clifford Behrens 1990 KAES: An Expert System for the Algebraic Analysis of Kinship Terminologies. Journal of Quantitative Anthropology 2:353-393.
READ, Dwight W., and Nicholas Gessler 1996 Cyberculture. In Encyclopedia of Cultural Anthropology. Human Relations Area File. M. Ember, ed. New York: Henry Holt.
VITZTHUM, Virginia J. 1994 Comparative Study of Breastfeeding Structure and Its Relation to Human Reproductive Ecology. Yearbook of Physical Anthropology 37:307-349.
WOOD, James 1994 Dynamics of Human Reproduction: Biology, Biometry, Demography. New York: Aldine de Gruyter.
 Return to Contents of this issue
Return to Contents of this issue
© Copyright Journal of Artificial Societies and Social Simulation, 1998
{kind=link}
{kind=link}
{kind=link}