
Introduction
Sexuality has been viewed as the nemesis of morality in nearly every traditional culture, yet in previous literatures in biology and genetics, sexuality is considered as one of the driving forces for the evolution of prosocial behaviours (Miller 2007). Selection in relation to sex provides a unique perspective to solve the persistent puzzle of the existence of social cooperation and altruistic behaviour (the strategy that benefits others and costs the player) that is referred to as the social dilemma (Dawes & Messick 2000). In other fields of virtues, biological evolutionary models showed that inter-sexual selection can explain the evolution of male-only care even if sexual selection may disfavour parental care when care comes at a net loss to fertilization success (Alonzo 2012), and provided templates for the virtue of altruism.
To solve the puzzle of cooperation, researchers proposed that altruism can benefit the fitness of the cooperator’s genes (Preston 2013). Based on this proposal, biologists have investigated in ways of inclusive fitness (Hamilton 1964; Dugatkin 2007) and various selections at different levels (Alexander 1987; Boyd and Richerson 1990; Sober & Wilson 1999; McAndrew 2002). Particularly, pioneering works have attempted to explain the maintenance and evolution of kin altruism (Hamilton 1964) and reciprocal altruism (Trivers 1971; Ridley 1997; Axelrod & Hamilton 1981; Sugiyama et al. 2002) via various forms of game theory, social selection models and empirical studies. In the field of pure altruism, the introduction of a costly altruistic punishment found in economic experiments (Fehr & Gächter 2002; Gintis 2000) paves the way for the emergence of altruism involving no genetic or direct rewards.
A costly punishment from a cooperator is not an advantageous strategy in the game of life, given other cooperators’ non-punishment strategy (Ye et al. 2011). This violation of causality, which is also referred to as the second-order social dilemma (2007), was partially solved by Hauert et al. (2007) via a dynamic evolutionary model where players could opt out of the collective action. The possibility of playing the nonparticipant strategy requires crucial assumptions concerning human society. Otherwise, the model loses its power to provide a cogent mechanism for the establishment of punishment (Boyd & Mathew 2007). To expand the breadth of the ‘narrow way to cooperation’ proposed by Hauert et al., various adjusted model and simulations have been conducted without the assumption of nonparticipation. Jaffe and Zaballa (2010) proposed a new form of punishment called cooperative punishment that was contingent upon the concurrent participation of other players. With the existence of cooperative punishment, social cohesion might be achieved in computer simulations. Hang Ye et al. (2011) introduced the moral concept of sympathy as compensation for the punisher. In Ye’s model of punishment and sympathy, individuals in the society are classified into three types: cooperator, defector and punisher. They share the benefits from public goods equally while only defectors do not contribute. Punishers impose an altruistic punishment on defectors at a cost to themselves, and cooperators who do not punish the defectors directly sympathize with the punisher and bear a cost due to the sympathy. The existence of sympathy in the model operates as a stabilizer for the social cohesion, where the punishers will persistently dominate the population.
All the adjusted models based on altruistic punishment are contributive and salient. However, taken alone they do not elucidate an understanding of how sexuality shapes the evolutionary equilibrium of altruism in human society. Besides, theoretical biologists and economists have broadly studied the benefit-cost analysis of altruism (Axelrod & Hamilton 1981; Axelrod 1984) without exploring the human motivation for selfless help (Preston 2013), while sexuality possibly plays the role of motivation to invest in altruism according to sexual selection theory and costly signalling theory (Gintis & Smith 2001; McAndrew 2002). The ignorance of sexuality in models involving alternations of generations is usually associated with the implicit assumption of asexual reproduction, which is a tradition from the Moran process (Moran 1958). As has been mentioned before and noted by many, sexual selection is a key factor in the evolution of human altruism. Without the analysis of the effect of sexual selection in altruism, the models are not complete for the simulations of human societies.
In this paper, we propose a dynamic model of punishment and sexual selection, where the social cohesion can be achieved by the behaviour-based sexual attractiveness from males that will increase the fitness of male punishers. Sexual attractiveness via sexual selection is an analogous to the sympathy in Ye’s model (Ye et al. 2011). The fundamental difference between Ye’s sympathy and sexual attractiveness in this paper is the division between asexual and sexual reproduction. The concept of sexual attractiveness in the model opens up the novel sources to study the game of public goods, altruism and other prosocial behaviours from the perspective of sexuality.
Baseline model
Model setup
We model the dynamic evolution of an isolated population of a constant scale N (we will modify this constant population assumption in next sections). The population is composed of four types of people including:
- MP: Male Punisher
- MD: Male Defector
- FC: Female Cooperator
- FD: Female Defector
The strategies and behaviours of each type are described as follows:
- MP is altruistic; that is, he will invest I into the altruistic behaviour. The investment incurs a cost c, which is assumed to be proportional to the investment (i.e., c = αI). However, MP also punishes defectors regardless of their genders. The strength of the punishment is δ ≥ c, which will incur a cost of γ < δ to MP. The population of MP is assumed to be NMP.
- MD is egoistic, contributes nothing to the altruistic behaviour and hence bears no altruistic cost. However, as a member of the society, he benefits from MP and FC’s contribution just. MD suffers punishments from MP. The population of MD is NMD
- FC is altruistic. She also invests I into the altruistic behaviour (we assume that I is identical in every MP and FC, and this assumption will be generalised to individual variation in the later section). FC also admires MP with the strength of s (i.e., the strength of sexual attractiveness or MP’s sexual capital), while the cost incurred by the s is set to 0. The population of FC is NFC
- FD is the same as MD except for their genders. For simplicity, we assume that FD will not admire any MP. The population of FD is NFD.
Every individual equally shares the benefit from the altruistic contribution from MP and FC.
It should be noted that the names of 'defector' or 'cooperator' do not necessarily imply that the individuals are involved in a typical public goods game defined in the field of economics. It would be more appropriate to rename the individuals in the society as ‘altruist’ and ‘egoist’. However, to pay tribute to the previous contribution by pioneers, we utilize their system of names in this paper.
By the definition of birth rate by Koella (2000), the expected payoffs for each individual in each of the four types in a single period are defined as:
| $$ P_{\mathit{MP}}=\beta \left(1-e^{-k\widetilde I}\right)-\mathit{\alpha I}-\gamma\left(N_{\mathit{MD}}+N_{\mathit{FD}}\right)+sN_{\mathit{FC}} $$ | (1) |
| $$ P_{\mathit{MD}}=\beta \left(1-e^{-k\widetilde I}\right)-\delta N_{\mathit{MP}}$$ | (2) |
| $$P_{\mathit{FC}}=\beta \left(1-e^{-k\widetilde I}\right)-\mathit{\alpha I} $$ | (3) |
| $$P_{\mathit{FD}}=\beta \left(1-e^{-k\widetilde I}\right)-\delta N_{\mathit{MP}}$$ | (4) |
| $$\widetilde I=\frac{\sum I_i} N=\frac{\left(N_{\mathit{MP}+N_{\mathit{FC}}}\right)I} N$$ | (5) |
We adopt the exponential form for the function of fitness (i.e., f = eωP), where f denotes the fitness of an individual, P denotes the payoff and ω denotes the selection strength (0<ω<1). The form of fitness comes from Traulsen et al. (2008).
We adopt a generalised Moran Process adjusted for sexuality and sexual reproduction to model the evolution of the society given the assumptions above. The probability of death d is identical to each individual in the society and remains constant during each period. To keep the population unchanged, the number of newborn offsprings is set to be equal to the number of the dead in the period. The process of determining the type of the offspring is completed through two steps: (1) nature randomly decides whether the offspring is male/female with a probability of 50% respectively; (2) if the offspring is a male, then his probability of being an MP (MD) is proportional to the fitness of an MP (MD), and if the offspring is a female, then her probability of being an FC (FD) is proportional to the fitness of an FC (FD). This process permits the balance of both genders.
Additionally, we assume that any individual of the population could randomly switch to another type within the same gender with a small probability μ, which is referred to as the mutation rate.
Simulation Results
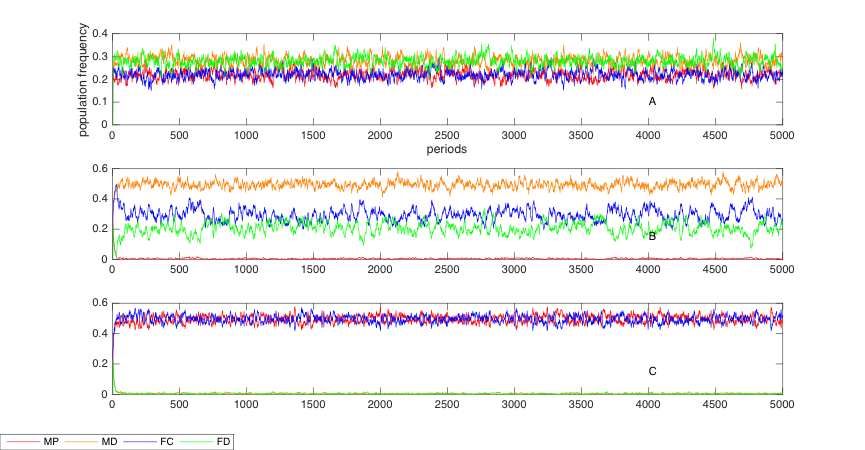
The outcome of the simulation is notable. Figure 1 shows that when punishment and sexual attractiveness exist (see Figure 1C), being altruistic is advantageous, which means that MP and FC are the dominant types in the society, whereas MD and FD are at the limit of near-extinction after some initial periods. However, in Figure 1A (no punishment and no attractiveness) and 1B (punishment without attractiveness), the status where the egoists become dominant over the altruists (especially in the male society) is a superlative example of the second-order social dilemma in a society with no sexual attractiveness. The difference in results between the simulations in Figure 1A, 1B and 1C indicates the effect of sexuality as the driving force to overcome the social dilemma of costly punishment. The apparent absence of sexual attractiveness and sexual selection in the evolutionary dynamics raises the extinction of punishers in Figure 1B. Meanwhile, the sexual attractiveness of a male punisher to a female cooperator shapes the evolutionary equilibrium where the virtue of altruism can escape the Darwinian Demon of ecological selection.
Robustness Check
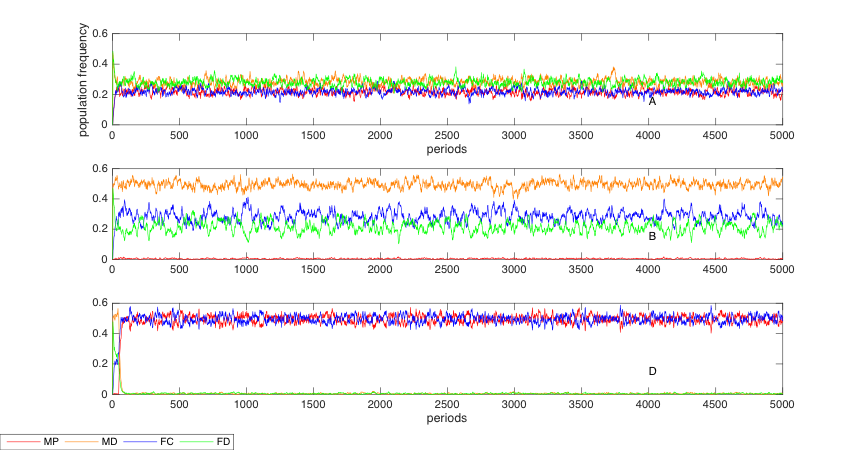
We perform robustness checks on various parameters and initial settings to test the power of our model. We select several important parameters to report their robust test results. The conclusion withstands the robust test on the initial proportion of the four types in the society. Figure 2 illustrates the results of the robust test with no MP and FC at period 0 with three different treatments as in Figure 1.
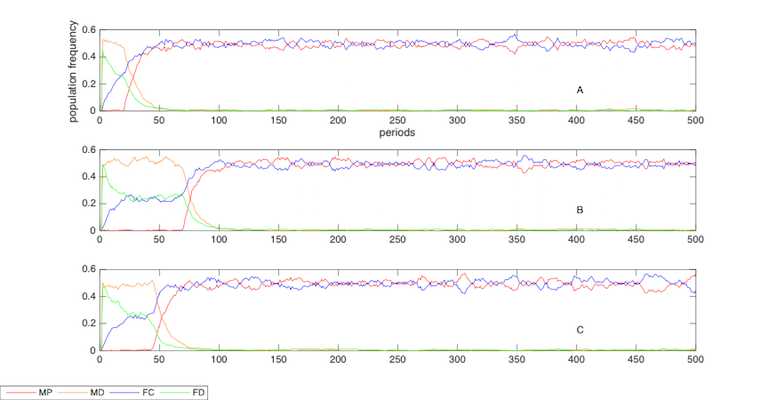
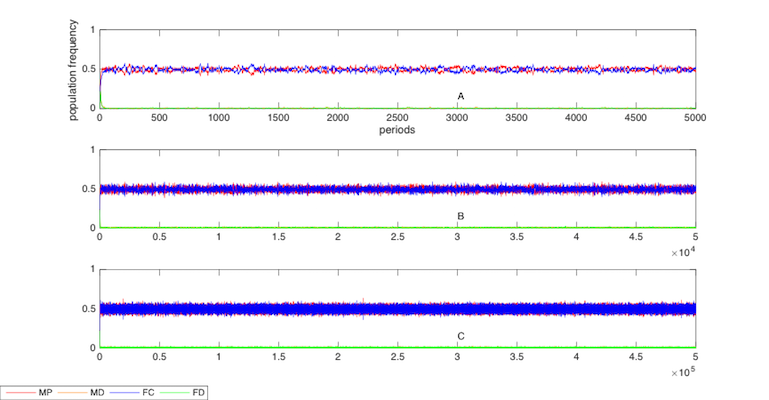
It is necessary to clarify that the exact timing of the turning point where altruists take power in the society from the egoists is inconsistent in repeated computer simulations due to the randomness of the model dynamics. Figure 3indicates the various timings of the turning points from three repeated computer simulations that report different evolutionary paths and routines.
We now conduct the robust test on the strength of sexual attractiveness, s.
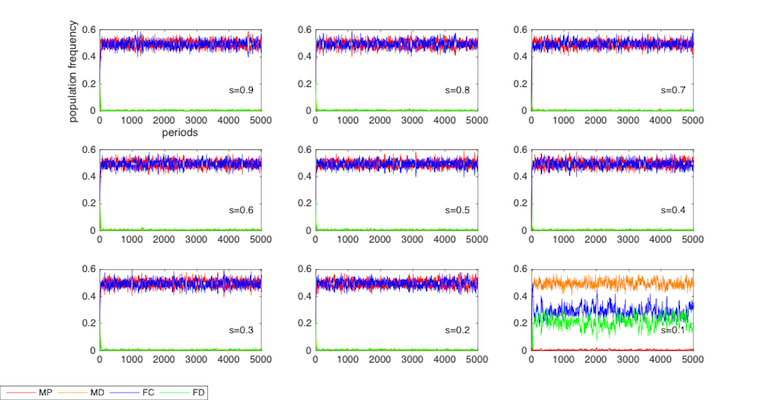
In simulations where s > 0.2, MP and FC take dominance over MD and FD; in the simulation where s = 0.1, MD takes dominance over MP while FC and FD coexist. The parameters in these simulations are the same as those in Figure 1C, with the exception of the value of s. It should be noted that the results for s = 1 and s = 0 have been reported in previous simulations.
Figure 5 tests the value of mutation rate μ, which is 0.001 in the baseline model.
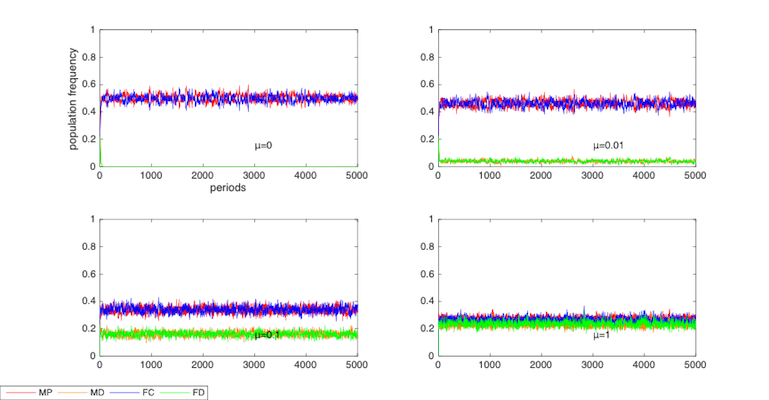
Figure 5 shows that mutation plays the role of ‘revolution’, trying to redistribute the population. A smaller mutation rate can widen the gap between cooperators and defectors.
Figure 6 shows the robust test for periods. The results withhold long periods.
Additionally, robust tests on other parameters are conducted to ensure the stability of the simulation results in this section.
Discussion of the Baseline Model
In Ye’s simulation model, human sympathy can be expressed by care, praise, solace, or even adoration and admiration (Ye et al. 2011). However, the moral sense of sympathy proposed and discussed by moral philosophers and moral psychologists cannot compensate the punisher directly by increasing the punisher’s fitness. Mate preference (or equivalently sexual attractiveness from the opposite perspective) serves as a particular form of sympathy that can increase the fitness of the individual to whom the preference is given.
The simulation results describing the evolutionary path of each type of individual provide critical evidence for sexual attractiveness as a strong stabilizer to generate a social equilibrium where prosocial strategy (i.e., being altruistic) is advantageous in an artificial human society. The introduction of sexuality to the traditional altruistic punishment game is beyond the scope of the previous literatures on social evolution via methods of computer simulation. We found that the introduction of sexuality and sexual attractiveness changed the social equilibrium from the sole dominance of punishers (Ye et al. 2011) to the coexistence of both (male) punishers and (female) cooperators, which fits reality better.
Extension to Variable Size Population
A strict constraint in Moran process is the constant population size, which greatly simplifies the calculations for theoretical modelling and serves as a common default in the field of evolutionary simulations. However, the assumption of fixed population will be fulfilled only in models with hard resource limitations (Claussen & Traulsen 2005), e.g. lattice models with viscous populations (Koella 2000). By assuming constant population, the interplay between reproductive strategies and population dynamics is neglected and only the effect of altruism on each individual’s fitness is taken into account. Even at capacity, the behaviour of altruism can increase the population size due to cooperators’ contribution to the public goods. The dependency of population size on cooperators happens when individuals in an isolated society compete for limited resources: egoism leads to the so-called common tragedy and altruism leads to abundant public goods due to the scale of economics (Houchmandzadeh 2015). Simulations have led to the outcome that an increasing population has a diminishing effect to produce a higher mean altruistic level in structured societies with a spatial distribution of agents (Shutters & Hales 2015). In Houchmandzadeh’s asexual model (2015), where population size is designed to be a function of the proportion of cooperators, clarified that a deleterious mutant (i.e., altruism) may possibly invade the community than the wild type (i.e., defectors). Based on this approach, it is reasonable to suppose that the variable size population (as a function of altruism) is an amplification factor for the social cohesion made by altruistic punishment and sexual selection as we have demonstrated in the preceding sections.
In this section, we apply the variable size population condition instead of the original constant population constraint to test the contribution of both sexual selection and population dynamics.
Variable Size Population Assumption
Consider an artificial society with four types of individuals as described before. Population size N is now an increasing function of the proportion of cooperators in the population (denoted as x), following Houchmandzdeh’s (2015) assumption. In each generation, the population reaches its capacity for the particular population structure (i.e. the distribution of each type), that is, no more individuals can survive under the resource limitations. To simplify the model, we adopt the linear form for N(x) (Houchmandzadeh 2015):
| $$ N\left(x\right)=\left(1-x\right)N_D+xN_C $$ | (6) |
| $$ x=\frac{(N_{\mathit{MP}}+N_{\mathit{FC}})} N $$ | (7) |
Simulation results
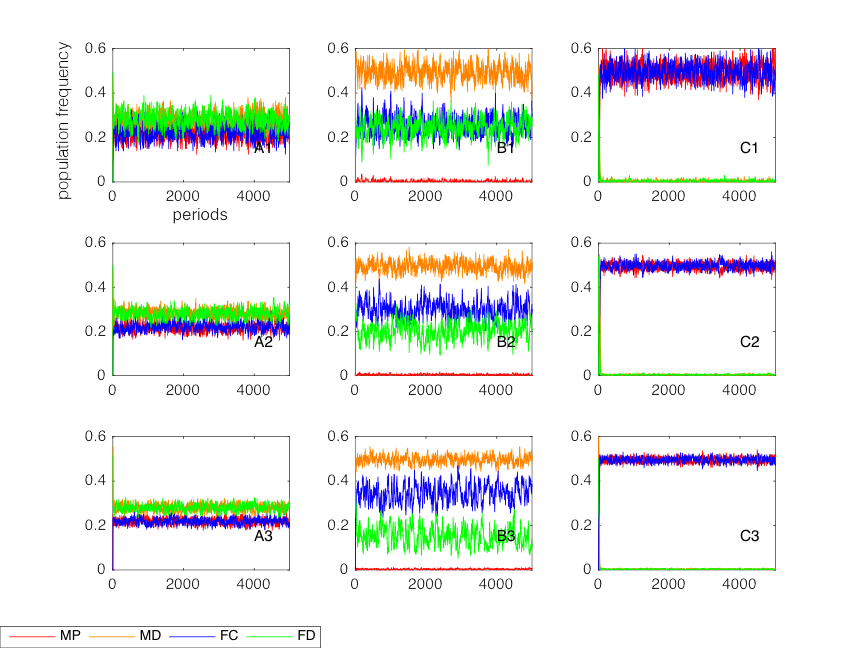
No remarkable differences in x from the baseline model can be detected when the initial population distribution was set to be evenly distributed between four types. To extract the invasion capacity of cooperators under variable size population assumption, we plot a comparison of simulation results for different treatments (Figure 7).
| Figure | Punishment** | Attractiveness | Pop. Dynamics*** | x at period 5,000 | Final population |
|---|---|---|---|---|---|
| A1 | (0,0) | s=0 | (200,200) | 0.4416 | 200 |
| B1 | (0.4,0.6) | s=0 | (200,200) | 0.2601 | 200 |
| C1 | (0.4,0.6) | s=1 | (200,200) | 0.9865 | 200 |
| A2 | (0,0) | s=0 | (1000,200) | 0.4392 | 551 |
| B2 | (0.4,0.6) | s=0 | (1000,200) | 0.3208 | 442 |
| C2 | (0.4,0.6) | s=1 | (2000,200) | 0.9904 | 992 |
| A3 | (0,0) | s=0 | (2000,200) | 0.4396 | 991 |
| A3 | (0.4,0.3) | s=0 | (2000,200) | 0.3501 | 830 |
| C3 | (0.4,0.6) | s=1 | (2000,200) | 0.9909 | 1983 |
The results of A1, A2 and A3 indicate that the differences in x are not significant, and the differences in x in C1, C2 and C3 are not significant as well. Now we compare the plots for FC and FD in B1, B2 and B3 to see the role of variable size population treatment in the change of invasion capacity. The value of x increases from B1 to B3. In B1, the plots of FC overlap with FD, that is, FC and FD share nearly the same population frequency (about 20%). When the variable size population treatment is employed (B2 and B3), the plots of FC and FD separate and FC gains the majority of females. The advantage of FC is enhanced in B3 compared to B2 due to a larger difference between NC and ND. Thus, in the society with altruistic punishment but without sexual attractiveness, variable size population treatment offers a more cooperative female society (i.e., FC dominates FD). In other words, FC in variable size population model has a higher invasion capacity than in constant population model with this particular setting. The result coincides partially with Houchmandzadeh’s theoretical outcomes in an asexual society.
Overall, these simulation results imply that the treatment of variable size population has an amplifying effect on the invasion of FC in the case of altruistic punishment model. Varying population size serves as a partial substitute for attractiveness in the altruistic evolution of females. The mechanism of variable size population in altruistic evolution is not clear, but it is clear that varying population size benefits the cooperative society as a whole: a defector-dominated society is designed to have less population size in the next period as a punishment. The increase in population induced by cooperators can overcome the altruistic cost (Houchmandzadeh 2015) and promote the rational consumption of limited resources.
Extension to Individual Variation
Variations in an individual’s social environment and state variables, such as colour, size and age, can influence the evolution of altruism, but the interaction between individual variation and cooperation have rarely been studied in an evolutionary way (Barta 2016). Therefore, an important problem in the evolutionary models based on the average population payoff functions, as the simulations we have done in the previous sections, is how to model the individual variation in the society. The study of penduline tits has been done based on the individual variation to resolve the conflict over parental care (Van Dijk et al. 2011). In the baseline simulation models, we classify the society into four types (i.e., MP, MD, FC, FD) for the sake of simplicity, but it is reasonable to question the accuracy of the model since a well-fitted simulation requires more than four types of individuals. We believe that in the real world, if altruistic investment can be scaled, the value of I follows a certain distribution and thus there are no strict stereotypes as punishers, cooperators or defectors.
In the extended models in this section, the society is composed of males and females investing I into altruism. I takes value from 0 to 1, with a random mutation changing the I in steps of ±0.1 (the settings are adopted from Koella’s (2000)) with a constant and identical mutation rate μ. Initially, altruistic investments in males and females both follow an identical normal distribution with a mean of 0.5 and standard deviation of 0.1.
The strategies and behaviours of each individual are assumed to satisfy:
- For male investing I* into altruism, he will punish individuals whose I <I* regardless of his/her gender. In turn, he will suffer from males whose I>I*
- For females investing I**into altruism, she will suffer from males whose I>I**. She will favour males whose I>I**.
To summary, a male will punish individuals with a lower level of altruism than him, and a female will favour males with more altruistic than her (i.e., a male favoured by a female will obtain a unit of sexual attractiveness s). Note that only males punish, only females show favour to males and only males receive female’s favour and thus increase their sexual attractiveness
The dynamic process is the same as in previous sections except the second step of birth. In this section, if the offspring is a male, his probability of having I=I* is
| $$ prob(I^{\ast} | male) = \sum f(I^{\ast} | male) / \sum f(I | male) $$ | (8) |
For the simulation results, we report the line graphs of the average value of I of males and females respectively. Figure 8 examines the effect of punishment and attractiveness on the mean value of I. Note that models settings in Figure 8 are listed in Table 2.
| Figure | Population Nmale/Nfemale | Period | Punishment (γ,δ) | Attractivenesss |
|---|---|---|---|---|
| A | 200/200 | 5000 | (0,0) | 0 |
| B | 200/200 | 5000 | (0.4,0.6) | 0 |
| C | 200/200 | 5000 | (0.4,0.6) | 1 |
| D | 200/200 | 5000 | (0.4,0.6) | 5 |
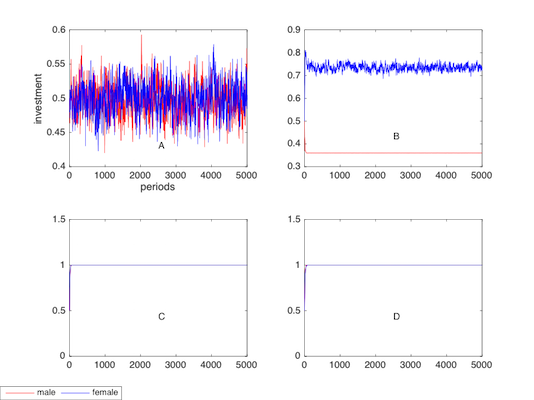
The plots in Figure 8 (B) show that altruistic punishment favours the evolution of high altruistic level females (I< 0.7) and disfavours the evolution of high altruistic level males (I<0.4). Nearly no fluctuations are observed in the plot of males in B. In Figure 8 (C) and Figure 8 (D), we employ both punishment and attractiveness (s=1 and 5 respectively). Plots in both C and D reach the level where I=1 regardless of genders.
To clarify, note that the mean value of I is a measure of the social altruistic level. A cooperative society will express a high mean value of I. Figure 8 (A)- Figure 8 (D) once again show the role of punishment and attractiveness in the altruistic evolution.
Finally, we report the final section (at period 5,000) of the investment distribution in Figure 9.
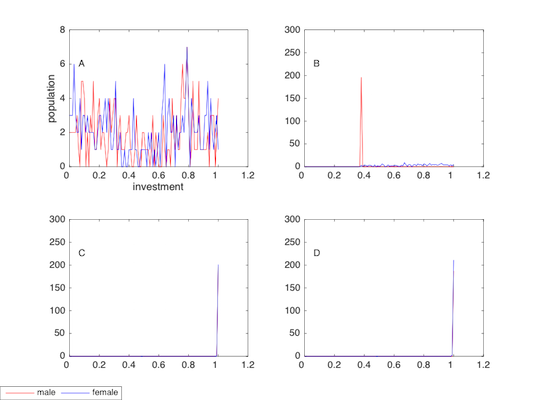
Given the normal distribution initially, individual’s investments are decentralised from the midpoint (I=0.5, the mean value of the normal distribution) in A. In B, most males invest about 0.4, and most females invest between 0.4 and 1, with a mean value larger than 0.7. In C and D, the plots of males completely overlap with females, and more than 200 males (females) invest 1, making them saints of their generation
In all, the results in this section are quite similar to the baseline model. The treatment of individual variation appears to be a significant amplifier for the evolution of altruism. Moreover, a superlative cohesion of society will be established where everyone invests 1 when both punishment and sexual attractiveness are employed. Surprisingly, the societies in C and D are even more cooperative than the cohesion achieved in the baseline model.
The above simulation results suggest that the evolution of altruism with individual variation assumption is a self-reinforcing process. The altruism in the society may be reinforced because the punishment mechanism runs synchronously with the evolution of altruism. Punishment and sexual attractiveness permit the advantage of cooperators, and the society structure may, in turn, enhance the punishment and attractiveness. For example, in simulation in Figure 8 (B), a male with I=0.8 suffers nearly no punishment and receives the highest honour of attractiveness at early period (i.e., period less than 10), but at later period (say 1,000 or more) he will suffer punishment from nearly every males and receive nearly no admiration if his investment remains 0.8 because the majority invest 1 now. The case for females is somehow similar. Our conclusion is quite optimistic in the sense that our society is likely to coordinate on high-level altruism equilibrium.
Discussion
In the baseline model and the extended models, the role of sexual attractiveness, variable size population and individual variation in the evolution of altruism are present in simulation results. In this paper, we provide a different and sexual approach to the puzzle of altruism, extending the existing evolutionary models in asexual evolutions. Based on the hypothesis that cooperation might be a costly commitment to public interest (Bird et al. 2012), we speculate that altruistic punishment is highly related to the sense of justice and law enforcement, and sexual attractiveness can be one of the predecessors of morality in civilisations.
Feminists may argue that the model setups in this paper suggest inequality between genders because only males are set to be qualified as punishers. Actually, females in this model can punish MD by not selecting them as sexual partners, which will lead to the corresponding effects of altruistic punishment on MD. Thus, in a generalised perspective, altruistic punishment is conducted by both MP and FC in the proposed model. Additionally, altruistic punishment can be performed in various forms: even uninvolved bystanders can pay to punish defectors to benefit the group (Raihani et al. 2010). Our simple assumptions on the gender-based behaviours are based on the historical observations, and for simplicity we do not model the effects of female punishers and male cooperators (who admire female cooperators/punishers). In modern civilisations, altruistic punishment is generally conducted by social norms and authorities, while mate preference to males/females (especially to males), such as superheroes and super heroines, with a strong sense of justice still exists.
Yet we have not talked about the incentives for human beings to value altruism as a sexually attractive trait, but it is hard to deny the fact that altruism, as well as other moral virtues, can inspire long-term love and admiration (Miller 2007). Researches in biology suggest that moral traits may serve as indicators of genetic quality and health condition in the process of mate choice (Neff & Pitcher 2005; Miller 2007). The hypothesis is that altruism evolves as a costly but honest signal to advertise the altruist in competition with others (Gintis et al. 2001). Religions and social norms also reinforce the mate preference to altruism by virtue of education and social scrutiny, and in turn provide infrastructures for the evolution of altruism through the mate preference. Religions also take part in creating of social cohesion directly via inhibition against norm violation ( Potgieter 2011). The mate preference we illustrate in this paper is not the only origin of the social cohesion because religions, social structures and politics also shape the society.
Finally, we have to note that the concept of altruism is understood differently in various disciplines (Clavien & Chapuisat 2012). In biology, altruism is defined by the behaviour that increases the fitness of another individual while decreasing the actor’s own fitness (Bell 2012). In our model, MP and FC are considered altruistic because at a particular instance (e.g., period 100) they sacrifice their own fitness for the sake of the whole society, although MP receives admiration from FC, which may lead to a net increase in MP’s fitness. The possible growth in MP’s fitness does not suggest that MP is biologically egoistic because the agent in the simulated model is treated as an artificial life or artificial molecular (Langton 1986) with no self-consciousness (i.e., MP will never realise that it is in his own interest to offer public goods). In this sense, the admiration from FC to MP is a life-long hidden blessing of sexuality.
Acknowledgements
This work was supported by the National Social Science Fund, China (Grant number: 13AZD061, 15ZDB134). For useful suggestions, we acknowledge Chenfeng Tu, Yize Chen, Chuankai Cheng, Ke Zeng and anonymous referees. We are most grateful to Zhun Deng for assistance in MATLAB codes.Appendix
Algorithm pseudo code for the baseline modelInitialize
Read in parameters
For each period:
For each individual:
Mutant with probability μ.
nvest, punish or admire according to type.
Calculate payoff P.
Calculate fitness by f=e^ωP.
Removed at a given probability d.
End
Set the number of newborns equals the number of death
Determine the type of each newborn according to the distribution of fitness
End
References
ALEXANDER, R. D. (1987). The biology of moral systems. Transaction Publishers.
ALONZO, S. H. (2012). Sexual selection favours male parental care, when females can choose. Proceedings of the Royal Society of London B: Biological Sciences, 279(1734), 1784-1790.
AXELROD, R. (1984). The Evolution of Cooperation. Basic Book.
AXELROD, R., & Hamilton, W. D. (1981). The evolution of cooperation. Science, 211(4489), 1390-1396.
BARTA, Z. (2016). Individual variation behind the evolution of cooperation. Phil. Trans. R. Soc. B, 371(1687), 20150087. [doi:10.1098/rstb.2015.0087]
BELL, G. (2012). Selection: the mechanism of evolution. Springer Science & Business Media.
BIRD, R. B., Scelza, B., Bird, D. W., & Smith, E. A. (2012). The hierarchy of virtue: mutualism, altruism and signaling in Martu women's cooperative hunting. Evolution and human behavior, 33(1), 64-78. [doi:10.1016/j.evolhumbehav.2011.05.007]
BOYD, R., & Mathew, S. (2007). Behavior. A narrow road to cooperation. Science, 316(5833), 1858-1859.
BOYD, R., & Richerson, P. J. (1990). Group selection among alternative evolutionarily stable strategies. Journal of theoretical biology, 145(3), 331-342. [doi:10.1016/S0022-5193(05)80113-4]
CLAUSSEN, J. C., & Traulsen, A. (2005). Non-Gaussian fluctuations arising from finite populations: Exact results for the evolutionary Moran process. Physical review E, 71(2), 025101.
CLAVIEN, C., & Chapuisat, M. (2012). Altruism–A Philosophical Analysis. eLS. [doi:10.1002/9780470015902.a0003442.pub2]
DAWES, R. M., & Messick, D. M. (2000). Social dilemmas. International journal of psychology, 35(2), 111-116.
DUGATKIN, L. A. (2007). Inclusive fitness theory from Darwin to Hamilton. Genetics, 176(3), 1375-1380.
FEHR, E., & Gächter, S. (2002). Altruistic punishment in humans. Nature, 415(6868), 137-140.
GINTIS, H. (2000). Strong reciprocity and human sociality. Journal of theoretical biology, 206(2), 169-179. [doi:10.1006/jtbi.2000.2111]
GINTIS, H., Smith, E. A., & Bowles, S. (2001). Costly Signaling and Cooperation. Journal of theoretical biology, 213(1), 103-119.
HAMILTON, W. D. (1964). The genetical evolution of social behavior. Journal of theoretical biology, 7, ll.
HAUERT, C., Traulsen, A., Brandt, H., Nowak, M. A., & Sigmund, K. (2007). Via freedom to coercion: the emergence of costly punishment. Science, 316 (5833), 1905-1907.
HOUCHMANDZADEH, B. (2015). Fluctuation driven fixation of cooperative behavior. BioSystems, 127, 60-66. [doi:10.1016/j.biosystems.2014.11.006]
JAFFE, K., & Zaballa, L. (2010). Co-operative punishment cements social cohesion. Journal of Artificial Societies and Social Simulation, 13(3), 4: https://www.jasss.org/13/3/4.html.
KOELLA, J. C. (2000). The spatial spread of altruism versus the evolutionary response of egoists. Proceedings of the royal society of London B: biological sciences, 267(1456), 1979-1985. [doi:10.1098/rspb.2000.1239]
LANGTON, C. G. (1986). Studying artificial life with cellular automata. Physica D: nonlinear phenomena, 22(1), 120-149.
MCANDREW, F. T. (2002). New evolutionary perspectives on altruism: Multilevel-selection and costly-signaling theories. Current directions in psychological science, 11(2), 79-82. [doi:10.1111/1467-8721.00173]
MILLER, G. F. (2007). Sexual selection for moral virtues. The quarterly review of biology, 82(2), 97-125.
MORAN, P. A. P. (1958). Random processes in genetics. Mathematical proceedings of the Cambridge philosophical society, 54, 1, pp. 60-71. [doi:10.1017/S0305004100033193]
NEFF, B. D., & Pitcher, T. E. (2005). Genetic quality and sexual selection: an integrated framework for good genes and compatible genes. Molecular Ecology, 14(1), 19-38.
POTGIETER, F. J. (2011). Morality as substructure of social justice: religion in education as a case in point. South African journal of edition, 31(3), 394-406.
PRESTON, S. D. (2013). The origins of altruism in offspring care. Psychological bulletin, 139(6), 1305.
RAIHANI, N. J., Grutter, A. S., & Bshary, R. (2010). Punishers benefit from third-party punishment in fish. Science, 327(5962), 171-171. [doi:10.1126/science.1183068]
RIDLEY, M. (1997). The origins of virtue. Penguin UK.
SHUTTERS, S. T., & Hales, D. (2015). Altruism Displays a Harmonic Signature in Structured Societies. Journal of Artificial Societies and Social Simulation, 18(3) 2: https://www.jasss.org/18/3/2.html [doi:10.18564/jasss.2780]
SOBER, E., & Wilson, D. S. (1999). Unto others: The evolution and psychology of unselfish behavior. Harvard University Press.
SUGIYAMA, L. S., Tooby, J., & Cosmides, L. (2002). Cross-cultural evidence of cognitive adaptations for social exchange among the Shiwiar of Ecuadorian Amazonia. Proceedings of the national academy of sciences, 99(17), 11537-11542. [doi:10.1073/pnas.122352999]
TRAULSEN, A., Shoresh, N., & Nowak, M. A. (2008). Analytical results for individual and group selection of any intensity. Bulletin of mathematical biology, 70(5), 1410-1424.
TRIVERS, R. L. (1971). The evolution of reciprocal altruism. Quarterly review of biology, 35-57. [doi:10.1086/406755]
VAN DIJK, R. E., Székely, T., Komdeur, J., Pogány, Á., Fawcett, T. W., & Weissing, F. J. (2011). Individual variation and the resolution of conflict over parental care in penduline tits. Proceedings of the royal society of London B: biological sciences, rspb20112297.
YE, H., Tan, F., Ding, M., Jia, Y., & Chen, Y. (2011). Attractiveness and punishment: evolution of cooperation in public goods game. Journal of Artificial Societies and Social Simulation, 14(4), 20: https://www.jasss.org/14/4/20.html. [doi:10.18564/jasss.1805]